|
 |
Pesci - Anatomia - Tessuti
|
|
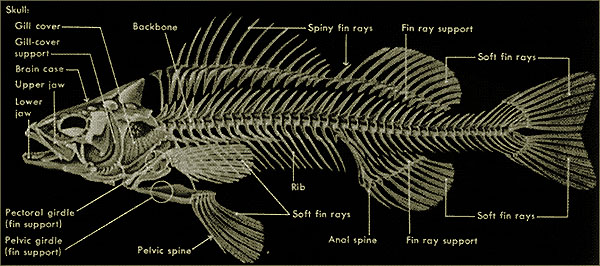
Tessuto Osseo - Scheletro |
Argomento trattato anche in
Apparato scheletrico
Tessuto scheletrico
Alle varietà dei tessuti che compongono le scaglie, i denticoli
cutanei e i denti, si deve aggiungere, nei pesci, la varietà non meno grande dei
loro tessuti endoscheletrici.
Il tessuto cordale assume presso i pesci
un'importanza che non si riscontra in nessun’altra specie.
I tessuti condroidi e
osteodi servono di passaggio tra le categorie abituali del connettivo, della
cartilagine e dell'osso che sono ben separati nei vertebrati superiori.
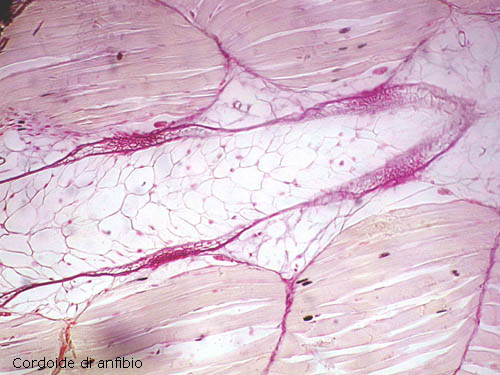
(Cordoide: tessuto di sostegno. Costituisce la corda dorsale (notocorda), presente durante
lo sviluppo embrionale in tutti i Cordati. Nel
tessuto cordoide è praticamente assente la sostanza fondamentale; le cellule,
una accanto all'altra, sono voluminose e globose e risultano deformate per
reciproca compressione; il citoplasma è scarso e confinato alla periferia della
cellula assieme al nucleo. Le cellule contengono
una notevole quantità di glicogeno che attira acqua, determinando il
caratteristico turgore di queste cellule.
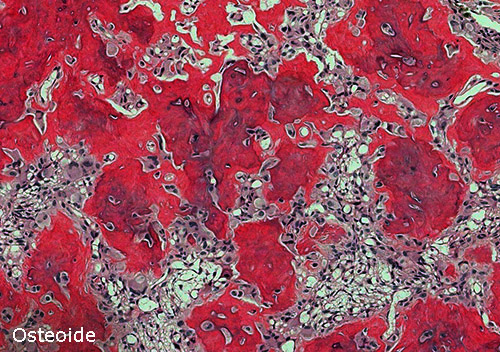
Osteoide: tessuto osseo povero di sali di calcio, costituito da
fibrille collagene che tendono a fondersi lateralmente, con focolai di iniziale
deposizione di calcio.
È prodotto dall’attività degli osteoblasti, e costituisce il precursore del
tessuto osseo.)
Secondo i gruppi, l'evoluzione si arresta allo stadio cartilagineo (condroitti) o
prosegue nella via dell'ossificazione (osteoitti).
La cartilagine può
calcificarsi senza ossificarsi. Un'altra particolarità del tessuto scheletrico
dei pesci è d'avere una crescita quasi illimitata.
L'osteogenesi si continua
durante tutta la vita, con rallentamenti o interruzioni solo nei periodi di
sottoalimentazione
Tessuti non calcificati
Tessuto cordale Notocorda
La corda dorsale forma la maggior parte dello scheletro assiale
dei ciclostoma, dei disponi e dei condorrei, e solo in alcuni stadi dello
sviluppo in tutte le altre specie, dove degenera più o meno precocemente ed è
sostituita dal tessuto osseo della colonna vertebrale.
Il residuo della
notocorda nei vertebrati prende il nome di nucleus polposus.
È il solo
tessuto scheletrico che ha origine direttamente da un epitelio e che sia
composto di cellule vacuolari. Ha proprietà meccaniche intermedie tra la
cartilagine e il connettivo cellulare. Le sue cellule contengono vacuoli e sono turgescenti.
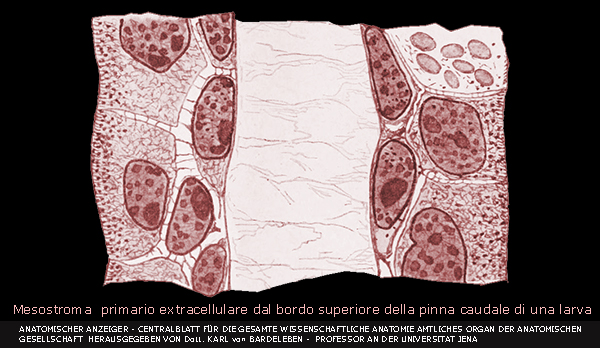
Mesostroma
Con la comparsa del mesoderma lo scheletro acquista dei nuovi
tessuti che sono raramente definitivi, ma servono da matrice alla cartilagine e
all'osso.
Il mesostroma è stato descritto come un insieme di filamenti
citoplasmatici che uniscono gli abbozzi degli organi e costituiscono dei ponti
intracellullari, che aumentano di numero e confluiscono in una rete spongiosa
dove mancano i nuclei cellulari ma dove compaiono poi delle fibrille finissime.
Forma lo scheletro gelatinoso o ialino negli embrioni nei pesci.
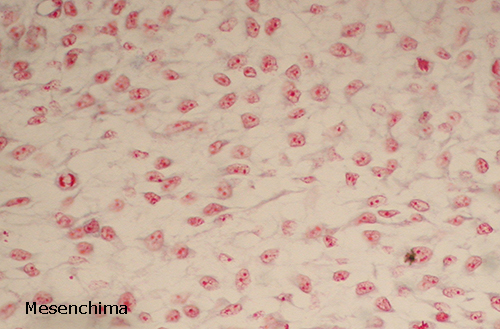
Mesenchima
Nella maggior parte dei pesci il mesostroma viene rapidamente
invaso dalle cellule mesenchimali, che derivano dal mesoderma e più precisamente
dalla parete splanica dei miotomi. Durante il periodo leptocefalico le cellule
mesenchimali restano alla periferia del mesostroma. Solamente nel corso della
metamorfosi lo scheletro gelatinoso è invaso dalle cellule periferiche e diventa
il manicotto schelettogeno che darà origine alle vertebre.
Il manicotto include la corda dorsale, il tubo neurale e i grossi vasi del tronco e della coda.
Il passaggio dal mesenchima a cartilagine si attua tramite un tessuto trovato in
tutti i pesci, formato da grandi cellule ovoidali, compatte, con nucleo piccolo
e citoplasma semiliquido, ialino e attraversato da tono-fibrille. Nelle cellule
si trova una minima quantità di sostanza fondamentale assai simile a quella
della cartilagine.
Cartilagine a stroma capsulare
È la sola rappresentata nei ciclostomi e si riscontra qua e là
anche nei selaci e nei teleosti (es.archi branchiali dove con l'età si trasforma
in ialina). la sua sostanza fondamentale è ricca di fibre elastiche (ramificate
isolate o in gruppi, la loro abbondanza è legata all'attività meccanica della
cartilagine in cui si trovano) e collagene (ondulate non ramificate).
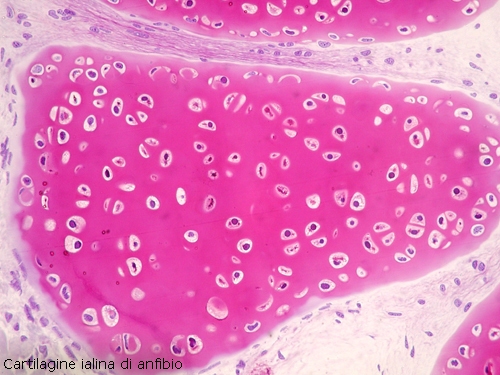
Cartilagine ialina
In questa varietà i condriociti diminuiscono di numero a
vantaggio della sostanza fondamentale che è traslucida, omogenea con un elevato
tenore in acqua e di condromucoide, contiene fibre collagene disposte su due
strati ortogonali tra loro.
Questa disposizione presente in molte varietà di
pesci e in molte parti dello scheletro, si estende dalla cartilagine fin nel
pericondrio e lo attraversa fino a fondersi al connettivo delle fasce muscolari.
I muscoli esercitano una forza sulla lamina pericondrale più prossima e quindi
sull’intero sistema di fibre e, dato che questi è presente in tutta la struttura
cartilaginea, il muscolo esercita la sua forza anche alla superficie pericondrale
lontana ed in ogni altro sito della cartilagine.
Questo sistema sembra prevenire
le fratture a livello cartilagineo. La cartilagine ialina è presente in certi
stadi dello sviluppo dello scheletro come anche in porzioni definitive formerà
lo scheletro transitorio o definitivo dei pesci.
Tessuti calcificati
Cartilagine calcificata
Quando ha luogo, la calcificazione della cartilagine consiste
in un deposito di materia minerale (carbonato e fosfato di calcio) nella
sostanza interstiziale e talvolta anche all'interno dei condriociti. Lo
scheletro dei selaci, di origine mesodermica è composto di: cartilagine ialina
nella cui sostanza fondamentale sono precipitati i sali di calcio.
Tessuto misto
Gli archi branchiali di alcuni teleosti (salmo, esox, gadus)
anch’essi di origine mesodermica, sono composti di cartilagine ialina, dove la
sostanza fondamentale periferica si calcifica e contemporaneamente le cellule
diventano fusiformi e parallele.
Tessuto osteoide
Compone lo scheletro dei teleostei superiori, l’origine è
sempre mesodermica: la sostanza fondamentale è ben calcificata e disposta in
lamelle, ma le cellule osteogeniche (osteoblasti) non sono ramificate e restano
eternamente al tessuto.
La calcificazione è sovente incompleta e la
mineralizzazione è influenzata dalle condizioni ambientali e dall'attività degli
organi. Il tessuto osteoide sembra essere meno resistente a infezioni batteriche
e protozoarie del tessuto osseo propriamente detto.
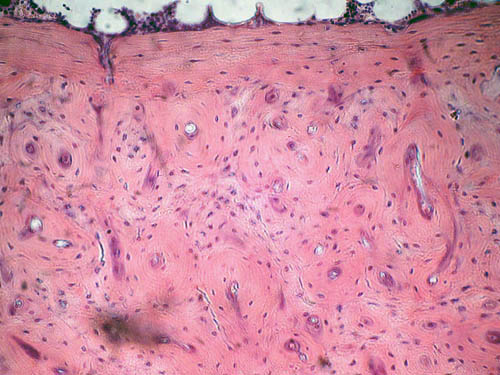
Tessuto osseo
Presente nello scheletro intero o in parte dei dipnoi, chondrostei e teleostei inferiori, è stato osservato anche negli archi
branchiali dei salmonidi, siluridi, ciprinidi e anguillidi adulti, l’origine
mesodermica, le sue componenti sono: osteociti ramificati e incorporati nella
sostanza fondamentale calcificata con rari canali di Havers
Tessuto osseo canalicolare
Compone lo scheletro degli holeosti, d’origine mesodermica, ha
cellule incluse (osteociti) ed esterne (osteoblasti) queste con dei
prolungamenti citoplasmatici che attraversano la sostanza fondamentale.
Evoluzione dei tessuti scheletrici
Condrogenesi
Tutte le cartilagini derivano dalla
trasformazione di un tessuto connettivo dove le cellule diventano
progressivamente dei condriociti e la sostanza fondamentale acquisisce proprietà
chimiche e fisiche nuove.
È molto più complessa per i particolari
rapporti tra tessuto osseo connettivo e cartilagineo .
Ossificazione acondrale
È l'ossificazione avviene in
assenza di cartilagine in seno a un tessuto connettivo fibroso come il
derma (ossa della volta cranica) o il manicotto schelettogeno (vertebre)
Ossificazione paracondrale
In questo caso l'osso si
costituisce in seno a un tessuto connettivo, ma nelle immediate vicinanze di
una placca cartilaginea (ossa del cranio che si sviluppano attorno alle
capsule nasali, ottiche e otiche).
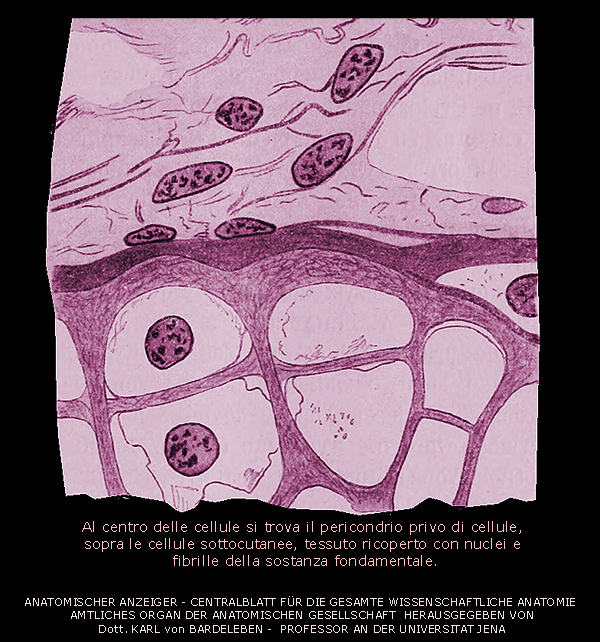
Ossificazione pericondrale
In questo caso,
l'ossificazione inizia a contatto con la cartilagine nel pericondrio
-
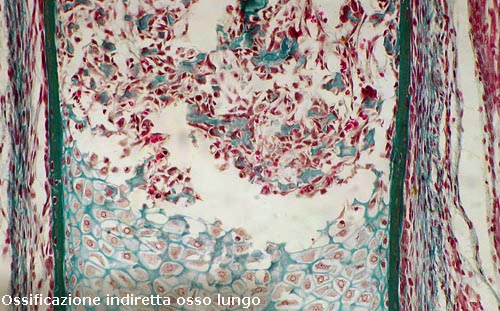
Ossificazione endocondrale
È la conseguenza e la
continuazione dell'ossificazione pericondrale, che inizia con un
rimodellamento della cartilagine dove le cellule s’ipertrofizzano e si mettono
in comunicazione tra loro favorite da una disidratazione seguita da una
ritrazione della sostanza fondamentale.
Si formano cavità irregolari; condriolisi e osteogenesi marciano alla pari.
Tutto lo spazio abbandonato
dalla cartilagine viene invaso da una neoformazione ossea.
L'ostogenesi nei pesci è illimitata, ma lenta, sovente tardiva
e s’interrompe sotto l'effetto di basse temperature e di un illuminazione
insufficiente
U. Fazzini
UniUD
2003-2004
AAVV
|