|
 |
Pesci -
Anatomia dei Pesci
|
|
Il Sistema Endocrino |
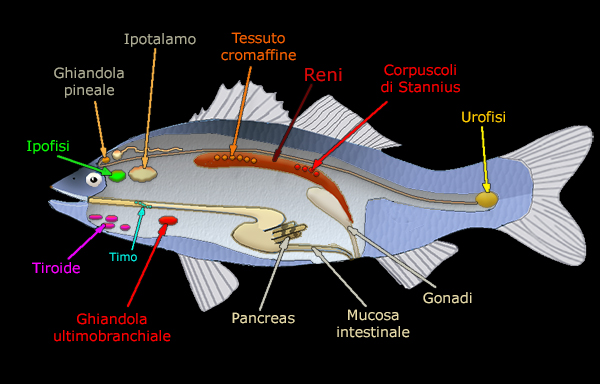
La
Ghiandola Pituitaria
(IPOFISI)
L'ipofisi dei
pesci č divisa in due parti:
I - L'adenoipofisi
II - Neuroipofisi
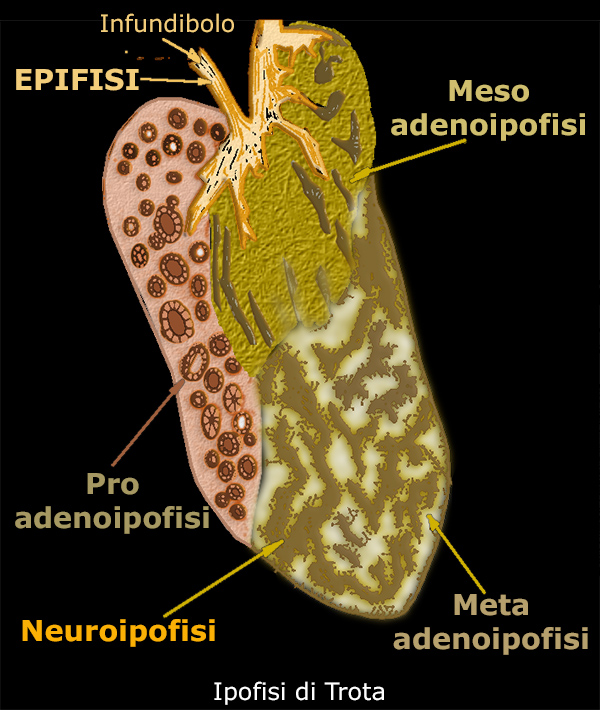
I -
Adenoipofisi
L'adenoipofisi
č suddivisa in tre parti.
A - pro-adenoipofisi.
B - meso-adenoipofisi.
C - meta-adenoipofisi
A)
Pro-adenoipofisi
Nella
proadenoipofisi si riconoscono tre tipi di cellule.
1 -
Cellule della prolattina
Nell'anguilla e nella trota iridea la maggior parte della
pro-adenoipofisi č costituita da cellule prolattiniche disposte in
modo follicolare. Le cellule prolattiniche sono acidofile e
contengono molti granuli secretori - 87 - colorabili con fucsina
acida o eritrosina. Tuttavia, questi granuli non si colorano con la
PAS (periodic acid-Schiff) o con la fucsina aldeidica (AF).
2 -
Corticotropi (cellule ACTH)
Queste cellule sono situate nella regione dorsale della
proadenoipofisi, formando due o tre strati al confine con la
neuroipofisi e contengono molti granuli secretori colorabili con
ematossilina di MacConaill. Tuttavia, le cellule ACTH non si
colorano con PAS o blu di Alcian.
3 - Tirotropi
(cellule TSH)
Nell'anguilla, le cellule TSH si trovano nella pro-adenoipofisi.
Sono macchiate tra i follicoli delle cellule prolattiniche e
talvolta appaiono come cordoni di cellule piů grandi. Nella maggior
parte delle altre specie di pesci, le cellule TSH si trovano
singolarmente o in gruppi nella mesoadenoipofisi. Queste cellule
sono basofile e contengono molti granuli secretori colorabili con
PAS, cromoematossilina (CH) o AF.
B)
Meso-adenoipofisi
Nella
meso-adenoipofisi si distinguono due tipi di cellule.
1)
Cellule dell'ormone della crescita (Somatotropi, cellule STH)
Le cellule STH occupano una porzione considerevole della
meso-adenoipofisi. Le cellule STH sono acidofile e non sono
colorabili con PAS o AF.
2) Gonadotropi
(cellule GTH)
Le cellule GTH non sono numerose nelle anguille con gonadi immature,
ma aumentano notevolmente nelle anguille mature. Talvolta invadono
la pro-adenoipofisi e vi si trovano nelle anguille e nelle trote
iridee con gonadi mature.
Le cellule GTH sono basofile e contengono molti granuli secretori
colorabili con PAS o AF.
C)
Meta-adenoipofisi
La
meta-adenoipofisi č composta da due tipi di cellule.
Un tipo non si colora con la PAS. L'altro tipo č PAS-positivo.
Queste cellule sono mescolate nella meta-adenoipofisi. Le cellule
PAS-negative sembrano secernere un ormone stimolante i melanofori
(MSH), che espande i granuli nei melanofori.
II -
Neuroipofisi
L'adenoipofisi č composta da tessuto ghiandolare, ma la neuroipofisi
č composta da tessuto nervoso contenente molte fibre nervose non
mielinizzate provenienti dall'ipotalamo e grandi cellule glia sparse
chiamate pituiciti.
La maggior parte delle fibre nervose contiene granuli secretori
colorabili con CH o AF.
I granuli sono chiamati
corpi di Herring
se vengono individuati come raggruppamenti nella neuroipofisi.
Nei pesci, la maggior parte di queste fibre nervose sono gli assoni
delle cellule neurosecretrici appartenenti al nucleo preottico
dell'ipotalamo.
I terminali di queste fibre nervose entrano in contatto con i
pituiciti o con le pareti dei capillari nella neuroipofisi o con le
cellule della metaadenoipofisi.
La neuroipofisi dei teleostei secerne due tipi di ormoni
ottapeptidici, la vasotocina arginina e l'isotocina (ittiotocina).
Sono prodotti nelle cellule neurosecretrici del nucleo preottico,
trasportati lungo l'assone e rilasciati nei capillari sanguigni
della neuroipofisi.
L'EPIFISI
L'epifisi č
composta da una
vescicola pineale e da un
peduncolo.
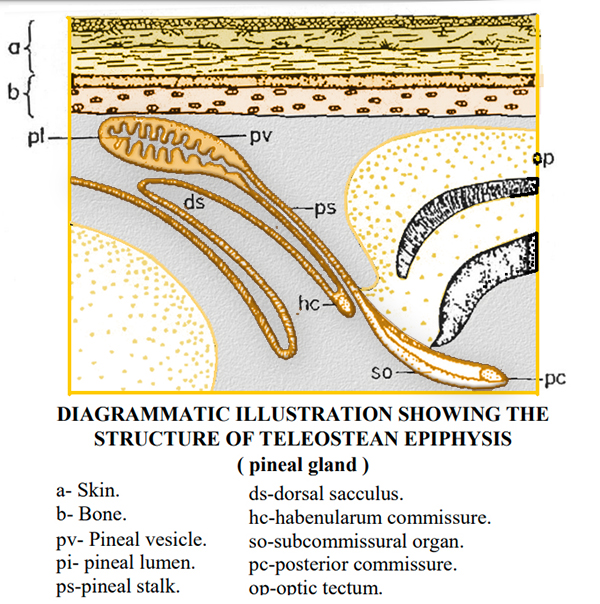
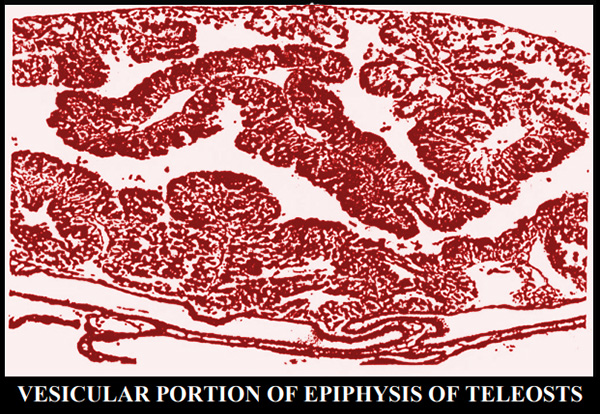
Il lume pineale della porzione vescicolare varia di dimensioni a
seconda delle specie. Il
parenchima epifisario č costituito da
cellule
fotorecettrici, di sostegno e gangliari, nonché dalla neuropila,
che č l'aggregazione dei processi citoplasmatici di queste cellule.
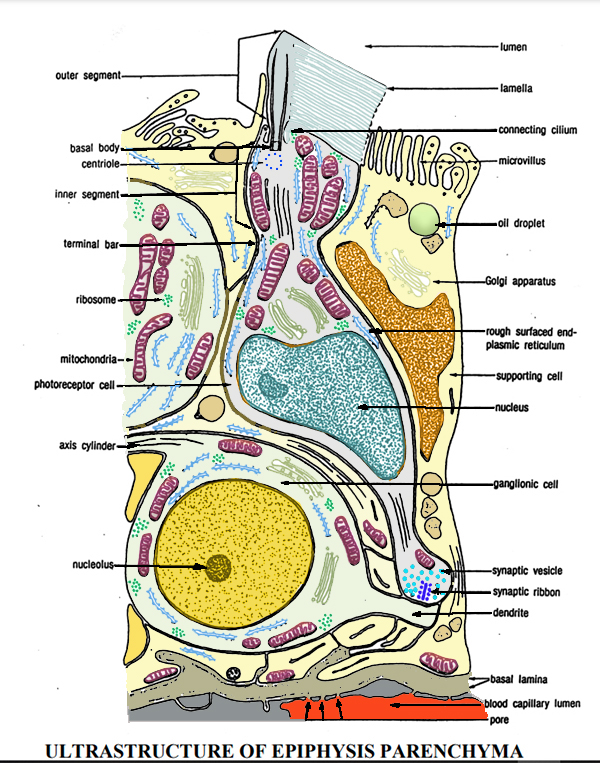
1) Le
cellule fotorecettrici
Le cellule
fotorecettrici dell'epifisi sono cellule sensoriali che presentano
fondamentalmente le stesse caratteristiche morfologiche delle
cellule del cono visivo della retina dell'occhio. L'osservazione al
microscopio elettronico rivela che la cellula fotorecettrice č
composta da segmenti interni ed esterni, corpo cellulare e processi
basali. I segmenti interni ed esterni sono costituiti
rispettivamente da mitocondri aggregati e da numerose lamelle.
2) Le
cellule gangliari
La presenza
di cellule gangliari nell'epifisi č stata confermata nella trota
iridea e nell'ayu, ma rispetto alle cellule fotorecettrici sono in
numero ridotto.
Le cellule ganglionari sono rilevabili nella regione centrale
dell'epitelio epifisario e, rispetto alle cellule fotorecettrici e
di supporto, sono di dimensioni maggiori.
Le cellule gangliari contengono nuclei grandi e prominenti, ma la
sostanza di Nissl nelle cellule non č abbondante.
Le cellule di sostegno riempiono gli spazi tra i fotorecettori e le
cellule gangliari.
3) Il
neuropilo
Le
osservazioni al microscopio elettronico hanno rivelato che questa
porzione č un'aggregazione dei processi basali di numerose cellule
fotorecettrici e dei dendriti delle cellule gangliari.
Questa aggregazione č chiamata neuropilo e qui i dendriti delle
cellule ganglionari formano sinapsi con i processi basali delle
cellule fotorecettrici.
Gli assoni non mielinizzati delle cellule ganglionari si riuniscono
e formano il tratto pineale, che si estende alla commissura
posteriore attraverso il peduncolo pineale.
L'UROFISI
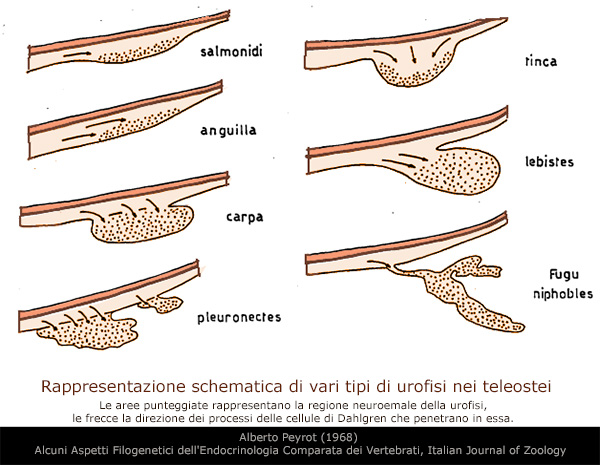
Nei teleostei, nella regione terminale del midollo spinale si
riconosce un piccolo corpo disteso chiamato urofisi.
Questo organo rilascia i materiali prodotti nelle cellule
neurosecretrici situate nel midollo spinale. Queste cellule,
insieme all'urofisi, sono chiamate sistema neurosecretorio
caudale.
Questo sistema neurosecretorio č riconosciuto solo nei teleostei e
negli elasmobranchi, ma č molto simile per organizzazione
strutturale al sistema ipotalamo-neurosecretorio presente nei
vertebrati.
Nel sistema neurosecretorio caudale, le cellule neurosecretrici sono
diffusamente localizzate nella regione terminale del midollo
spinale. I terminali degli assoni di queste cellule si riuniscono
sul lato ventrale della regione e formano l'urofisi con i capillari
sanguigni.
I terminali degli assoni terminano in corrispondenza delle pareti di
questi capillari nell'urofisi. La cellula neurosecretrice č una
grande cellula nervosa contenente un nucleo polimorfo e il suo
citoplasma č basofilo.
L'urofisi č composta da elementi del midollo spinale, come assoni
neurosecretori, glia e fibre ependimali e glia, e da derivati
meningei, come reticolo vascolare e fibre reticolari.
Nell'urofisi sono osservabili molti materiali neurosecretori
granulari. Questi materiali del sistema neurosecretivo caudale sono
colorati con azocarmina, ferro-ematossilina o floxina, che vengono
utilizzate come controcolorazione.
LA
GHIANDOLA TIROIDEA
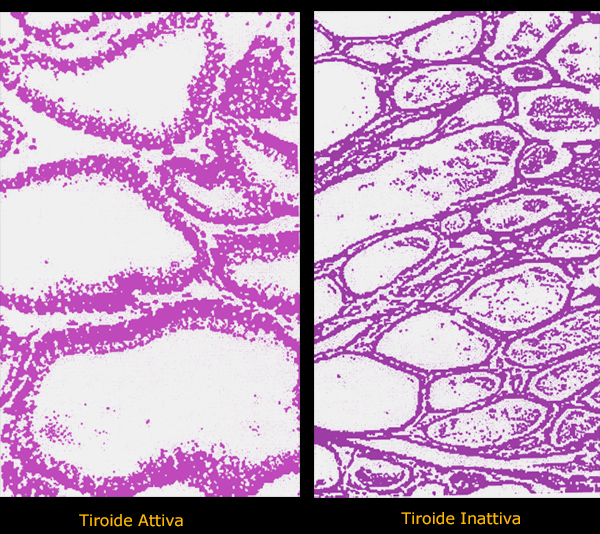
I follicoli
tiroidei di molti teleostei, a differenza di quelli di altri
vertebrati, si disperdono singolarmente o in gruppi nel tessuto
connettivo intorno alla parte divergente dell'arteria branchiale
afferente e non formano un organo compatto.
I follicoli sono di dimensioni variabili, rivestiti da epitelio
cuboidale semplice e riempiti di colloide nel lume. L'attivitŕ del
follicolo assorbe lo iodio dal sangue e produce gli ormoni tiroidei.
Quando la tiroide č attiva, le cellule epiteliali sono cuboidali e
c'č poca colloide. Per questo motivo si riconoscono spesso molti
vacuoli nella colloide adiacente all'epitelio. Quando invece la
ghiandola č inattiva, le cellule epiteliali sono squamose e il
lume follicolare č pieno di colloide.
IL TIMO
Il timo si trova sulla parete dorsolaterale della faringe, dalla
prima alla quarta arcata branchiale.
La sua superficie ventrale č
ricoperta da epitelio mucoso.
Sul lato dorsale č presente uno strato di tessuto connettivo e
successivamente uno strato di muscolo. Il parenchima č costituito da
tessuto linfoide, quasi interamente composto da linfociti.
Il timo č evidente negli avannotti, ma diventa meno prominente nei
pesci adulti.
Č stato riferito che in alcuni gobidi le dimensioni del timo
cambiano con il ciclo riproduttivo.
A seconda della specie, possono essere presenti strutture simili ai
corpuscoli di Hassal.
La localizzazione e
la struttura (non suddivisa
nettamente in zone corticali e midollari) del
timo variano da una specie ittica all’altra
Nei Teleostei, il timo č il principale organo linfoide e durante lo
sviluppo sembra infondere linfociti (forse
di tipo T) il rene e la milza.
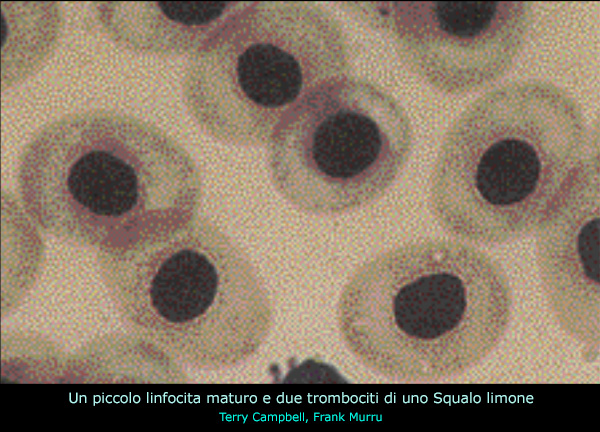
I linfociti sono i
leucociti piů comunemente riscontrati nel sangue periferico di molte
specie ittiche. Sono stati descritti
tre tipi di linfociti: maturi, immaturi (linfoblasti) e
plasmacellule.
I primi sono cellule piccole che tendono ad essere rotonde, ma
possono anche presentare un profilo irregolare o sagomarsi
adattandosi alla forma di quelle adiacenti.
Il significato della presenza dei linfociti immaturi nel
sangue periferico dei pesci č controverso. Alcune di queste cellule
possono rappresentare linfociti reattivi che stanno rispondendo ad
una stimolazione antigenica.
Le plasmacellule (che sono forme reattive di linfociti B) si
riscontrano occasionalmente nel sangue periferico dei pesci.
La maggior parte dei linfociti riscontrati nel sangue periferico dei
pesci č piccola e matura.
(Introduzione
alle ematologia dei pesci -
Terry
Campbell (Kansas State University),
Frank Murru (Sea World
of Florida) Florida)
LA GHIANDOLA
ULTIMOBRANCHIALE
La
ghiandola ultimobranchiale č chiamata anche
corpo soprapericardico
o ghiandola subesofagea.
Nella maggior parte dei teleostei, la ghiandola si trova nel setto
trasversale tra la cavitŕ pericardica e quella peritoneale, appena
sotto l'esofago.
La ghiandola ultimobranchiale dell'anguilla e della trota iridea č
costituita da una coppia di masse di tessuto a destra e a sinistra,
mentre la ghiandola della carpa č costituita da una sola massa.
Nella carpa il tessuto ultimobranchiale č ben sviluppato e composto
da molti follicoli densamente stipati. I nuclei delle cellule
ghiandolari sono rotondi o ovali e contengono poca cromatina. I
granuli secretori non sono visibili al microscopio ottico.
Nel tessuto ultimobranchiale dell'anguilla e della trota iridea si
osserva una cavitŕ centrale. Inoltre, nella ghiandola della trota
iridea si osservano pieghe che sporgono nella cavitŕ. In queste
specie, il parenchima della ghiandola ultimobranchiale non ha una
disposizione follicolare, ma č costituito da epiteli
pseudostratificati. Il nucleo č situato nella porzione basale del
citoplasma ed č di forma ovale. Nella trota iridea, le cellule a
calice sono presenti singolarmente o in gruppi tra le cellule
ultimo-branchiali.
LA
GHIANDOLA INTERRENALE E LE CELLULE CROMAFFINI
La
ghiandola interrenale dei pesci č omologa alla corteccia surrenale
dei mammiferi.
Di solito č distribuita nel rene anteriore e circonda la vena
postcardinale e i suoi rami.
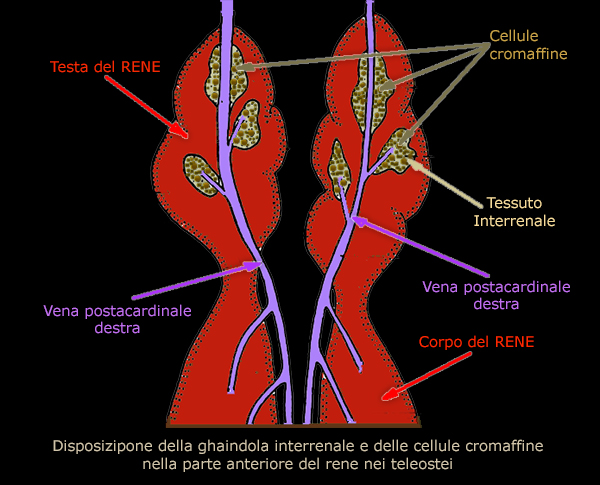
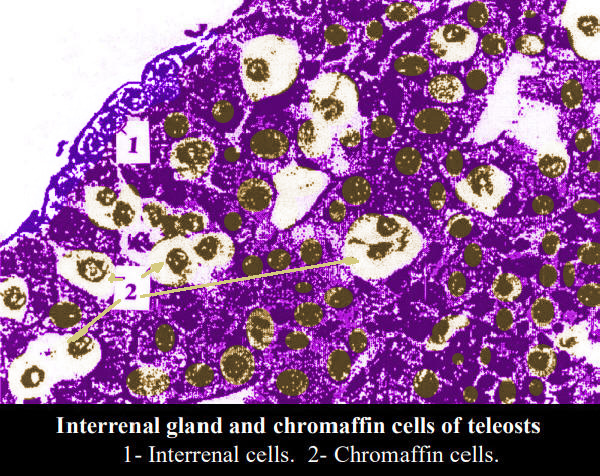
Nei pesci, il tessuto omologo alla midollare surrenale dei mammiferi
č chiamato cellule cromaffini e sono strettamente correlate
alla ghiandola interrenale.
L'identificazione delle cellule interrenali e cromaffini non č
difficile perché il citoplasma delle cellule interrenali si colora
profondamente con l'eosina.
Le cellule cromaffini sono quasi incolori nei preparati fissati nel
liquido di Bouin e colorati con ematossilina ed eosina. Se le
cellule cromaffini vengono fissate in un liquido contenente
bicromato, come il formolo di Zenker, e colorate con H-E, nel
citoplasma compaiono minuscoli granuli giallo-brunastri (reazione
cromaffina).
L'identificazione delle cellule cromaffini puň essere effettuata
facilmente grazie a questa reazione. Il nucleo della cellula
interrenale č solitamente rotondo, contiene poca cromatina e ha un
nucleolo distinto.
Il nucleo delle cellule cromaffini č di forma ovale o irregolare e
contiene anch'esso poca cromatina. Il nucleolo č oscurato nel nucleo
delle cromaffine. Di conseguenza, l'identificazione delle cellule
interrenali e cromaffini č possibile da queste caratteristiche dei
loro nuclei.
Le cellule interrenali dei teleostei sono colorabili con Sudan nero
B. L'esame istochimico delle cellule interrenali dei teleostei
rivela la presenza degli enzimi 3β-idrossisteroide deidrogenasi e
glucosio-6-fosfato deidrogenasi, importanti nella biosintesi degli
ormoni steroidei.
LE ISOLE
DI LANGERHANS
Le isole di Langerhans sono di origine endodermica e nella maggior parte dei
teleostei si trovano come piccoli corpi sparsi nella porzione
esocrina del pancreas.
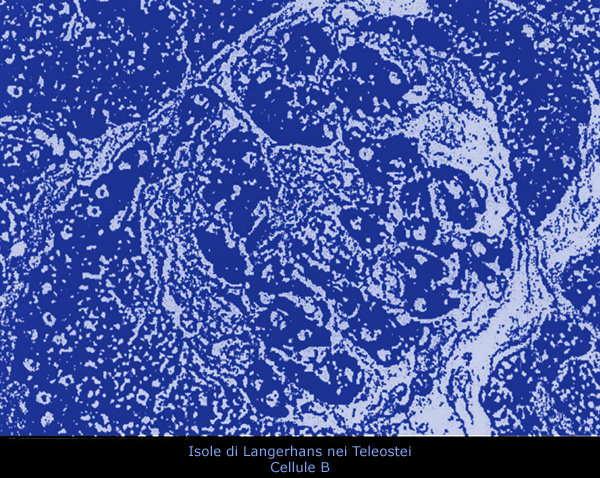
Nei teleostei, al microscopio ottico si possono distinguere tre tipi
di cellule nelle isole di Langerhans: cellule A, B e D.
1) Cellule A
Sono normalmente numerose alla periferia delle isole. Le
dimensioni e la forma di queste cellule variano notevolmente a
seconda della specie. In particolare nella carpa, presentano
dimensioni e forma irregolari.
Il nucleo presenta un'invaginazione
su un lato.
I granuli cellulari sono facilmente colorati con coloranti acidi
come la floxina e l'azocarmina se i campioni sono stati
precedentemente fissati in ZF. Tuttavia, altri fissativi non possono
ottenere risultati soddisfacenti.
Č stato dimostrato da molti ricercatori che le cellule A producono e
secernono glucagone, uno dei diversi ormoni che promuovono l'aumento
dei livelli di glucosio nel sangue.
2) Cellule B
Queste cellule producono, immagazzinano e secernono insulina, un
ormone ipoglicemizzante. Nelle isole piccole tendono a riunirsi al
centro, mentre nelle isole grandi sono disposte in cordoni e
mostrano un modello di distribuzione piuttosto complicato.
I granuli delle cellule B si colorano bene con AF e CH, quindi le
cellule B si distinguono facilmente dalle altre cellule
dell'isolotto.
In generale, le cellule B sono ovoidali o a forma di fuso e le loro
dimensioni sono simili o leggermente inferiori a quelle delle
cellule A.
3) Cellule D
Queste cellule si trovano spesso isolate tra le cellule B. La loro
forma č generalmente arrotondata o ovoidale, ma le loro dimensioni
mostrano una marcata variazione interspecifica. Sono piů piccole
delle cellule A e B.
I granuli delle cellule D sono leggermente colorati con floxina nei
campioni precedentemente fissati in formolo di Zenker, ma non nei
campioni trattati con altri fissativi. Questi granuli hanno forti
proprietŕ argirofile; pertanto le tecniche di colorazione con
argento sono utili per identificare le cellule D.
I
CORPUSCOLI DI STANNIUS
I
corpuscoli di Stannius sono piccoli corpi bianco-giallastri presenti
nella regione renale dei teleostei.
I corpuscoli di Stannius sono avvolti da tessuto connettivo
capsulare e il loro parenchima č diviso in molti lobuli. Ogni lobulo
č composto da molte cellule ghiandolari contenenti numerosi granuli
secretori grossolani nella porzione basale. Inoltre, nei lobuli di
alcuni pesci č possibile individuare una cavitŕ centrale o
pseudolume.
I granuli secretori del corpuscolo di Stannius sono profondamente
colorabili con eosina (colorazione HE), ferro-ematossilina
(colorazione di Heidenhain) e fucsina acida (tripla colorazione di
Mallory).
Nei corpuscoli di Stannius del salmone atlantico č stata rilevata la
presenza di ormoni steroidei come cortisolo, corticosterone e ormoni
sessuali.
Inoltre, nei corpuscoli di Stannius di anguilla europea, carpa e
rana pescatrice č stata trovata una sostanza simile alla renina.
Mohammad Rashad Fath El bab
Fundamentals of the HISTOLOGY OF FISH - PART
I
HISTOLOGY OF TELEOSTS
2006
|


















