|
|
 |
Pesci - Anatomia - Organi di senso
|
|
L'Orecchio
interno - Udito
|
I pesci
hanno un apparato acustico in grado di sentire i suoni compresi tra
16 e 5.600 Hz. Lĺorecchio interno e la linea laterale
sono gli organi che funzionano come un unico
sistema fondamentale per la recezione di stimoli pressori e
acustici dallĺambiente circostante. I
canali della linea laterale contengono meccanorecettori, noti come neuromasti, che costituiscono lĺunitÓ base del sistema. I neuromasti sono organi a forma di cupola costituiti da un gruppo
di cellule sensoriali cigliate generalmente con una cupola gelatinosa che include i fasci di
cilia che si estendono in alto al disopra dellĺepitelio. Le cellule sensoriali sono contornate da
cellule di sostegno e trasmettono gli stimoli al sistema nervoso.
L'Orecchio interno
L'orecchio interno, o labirinto membranoso, consiste in sacchi e canali che formano un sistema chiuso (contenente
un liquido detto endolinfa) e deriva da una vescicola otica dell'embrione - dĺorigine ectodermica (placoide
uditiva).
E' compreso nel labirinto cartilagineo scavato nelle regioni otiche del
neurocranio, dove Ŕ immerso un altro liquido detto perilinfa.
L'otocisti nell'embrione si compone in una parte pi¨ ampia detta
tasca
vestibolare e una pi¨ piccola detta tasca cocleare fra loro separate da
un restringimento che darÓ origine al labirinto composto di un
utricolo (tappezzato da neuromasti sensoriali su cui
poggiano delle piccole strutture ossee, gli otolitie
un) sacculo, che un restringimento separa dalla
macula della lagena
(vescicola
sede dell'udito, che corrisponde alla
chiocciola dei vertebrati terrestri), tre
canali semicircolari (due verticali perpendicolari tra loro e l'altro orizzontale).
Le cellule sensoriali che formano la macula assomigliano ai neuromasti che si
incontrano nella linea laterale. Queste cellule si collegano ai nervi acustici nel cervello.
Cristalli di carbonato di calcio sono depositati sopra la macula e si
combinano a formare gli otoliti.
Il sistema nervoso centrale interpreta questi segnali e
fornisce informazioni al pesce per il suo orientamento e movimento nello spazio.
Le ossa che delimitano la cavitÓ otica sono: il frontale, il parietale, lo sfenotico,
l'opistotico, l'occipitale.
Nei selaci la capsula otica Ŕ contenuta interamente nello spessore della cartilagine.
Selaci
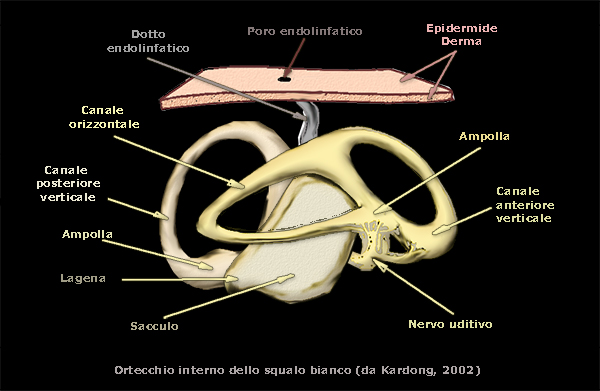
Orecchio interno negli squali con tre canali semicircolari, con otoliti, con funzione specialmente
statorecettrice,
ma con presenza anche di neuromasti con funzioni fonorecettrici Ŕ molto simile a quello dei mammiferi ed Ŕ
particolarmente sensibile ai suoni a bassa frequenza, mentre
lĺorecchio esterno Ŕ del tutto assente.
Lĺorecchio interno Ŕ connesso
allĺesterno mediante canali endolinfatici, i quali si aprono in due
piccoli forellini per lato sulla parte dorsale della testa.
Negli Elasmobranchi gli otoliti consistono di una matrice gelatinosa contenente numerosi piccoli cristalli di carbonato di calcio.
I tre canali semicircolari presentano delle disposizioni assai diverse.
Nel genere torpedo i canali verticali sono completamente indipendenti
l'uno dall'altro, ed Ŕ nell'anteriore che comunicano le estremitÓ del
canale semicircolare esterno (orizzontale).
In questo tipo di labirinto
si possono distinguere:
- una parte anteriore comprendente il canale semicircolare verticale anteriore, il
canale orizzontale e l'otricolo
-
una parte posteriore con il
canale verticale posteriore e il sacculo.
Presso gli altri selaci i due canali semicircolari verticali si
avvicinano al punto che le loro estremitÓ girate verso la zona interna
vengono a contatto tra loro e sono separate da una membrana (es:
leomargus) o possono fondersi insieme (es:chimera), che Ŕ la condizione
comune a tutti i canali semicircolari verticali dei vertebrati.
In ambedue i modelli di orecchio interno, dalla zona di congiunzione fra
utricolo e sacculo pare un canale verticale mediano: il
dotto linfatico,
che si apre nello spazio sottomeningeo periencefalico e all'esterno
sulla faccia dorsale del cranio, questi fa comunicare il labirinto con
l'esterno. Il dotto linfatico contiene un fluido gelatinoso in cui si possono osservare
delle particole solide provenienti dall'esterno.
Gli squali possiedono la capacitÓ di percepire dei cambiamenti di pressione
nell'orecchio interno e riescono a percepire suoni
tra 10 e 800 Hz e la macula negletta, consente loro di discriminare la
direzione dei suoni provenienti da lunghe distanze
Pesci non selaci
Il labirinto si presenta come nella chimera ma con un canale
endolinfatico in meno; i due rami dei canali semicircolari verticali
sono fusi insieme salvo nei pesci abissali. Le dimensioni relative dei
canali variano da un genere all'altro sono molto grandi nei mormyrili,
cyclopteri.
Il sacculo si separa in due compartimenti, uno, il pi¨
grande, il sacculo p.d. e l'altro la
lagena (questa
Ŕ ben delimitata nell'acipenser).
Il sacculo in certi pesci Ŕ molto
grande da superare in altezza l'arco superiore dei canali verticali (Ggobius
niger).
Manca il canale endolinfatico o persiste sotto forma di canale
cieco.
In alcuni ordini di Teleostei, tra cui i Siluriformes e i Cypriniformes (pesci ostariofisi), la parte cefalica
della vescica gassosa Ŕ collegata con il labirinto dell'orecchio interno attraverso l'apparato di Weber costituito da
una serie di piccole ossa derivate da parti delle prime vertebre. Questa interdipendenza funzionale tra
vescica gassosa e labirinto Ŕ finalizzata, oltre che alla regolazione dell'equilibrio e della posizione del pesce,
anche alla ricezione e alla produzione di suoni
(al di sotto di 3 kHz)
Diversi altri gruppi di teleostei possiedono modificazioni della
parte cefalica della vescica natatoria che interessano l'orecchio interno e possono influenzare la capacitÓ di ricevere ed emettere suoni
(in molti casi
i suoni prodotti anche dallo sfregamento di parti del
corpo tra di loro).
Struttura della parete del labirinto
Il rivestimento interno del labirinto Ŕ formato da un epitelio
pavimentoso semplice, si ispessisce localmente in alcune tipiche placche
sensitive, esternamente Ŕ rivestito da tessuto connettivo fibroso ben
vascolarizzato.
Le cellule sensoriali, nei canali semicircolari, si
localizzano nei rigonfiamenti detti ampolle del
tratto di origine dei tre canali, a causa del loro aspetto e della loro
localizzazione sono indicate con i nomi di
cresta
ampolliera anteriore, esterna o laterale e posteriore, le altre, pi¨' appiattite, sono chiamate
macule
o papille; si distinguono per la loro sede, una
macula dell'otricolo,
una del sacculo e una della
lagena e talora una macula negletta vicino al punto in cui dal sacculo si passa del canale semicircolare posteriore.
Le creste ampollose anteriore ed esterna e la macula dell'otricolo, appartenenti alla parte anteriore dell'orecchio interno, sono innervate
dal ramo anteriore del'VIII░ paio di nervi cranici.
Le macule del sacculo e della lagena, la macula negletta e la cresta ampollare
posteriore sono innervate dal ramo posteriore del medesimo nervo.
L'epitelio sensitivo di queste strutture, stimolato dai movimenti
dell'endolinfa, informa l'animale della posizione della sua testa nello spazio e dellĺentitÓ delle accelerazioni o decelerazioni che si
attuano durante il movimento sia della testa sia di tutto il corpo.
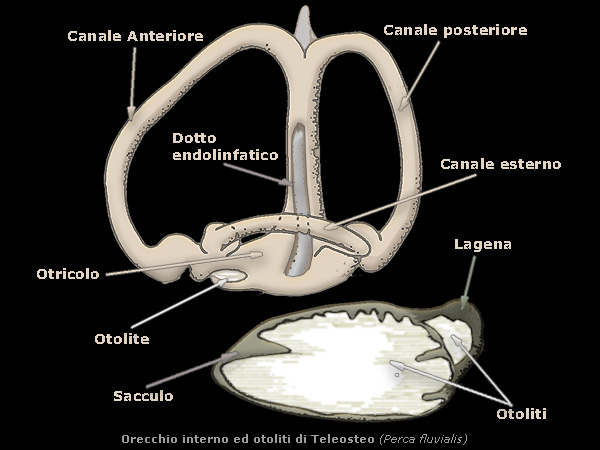
L'orecchio interno degli osteoitti ha un sacculo particolarmente sviluppato con annessa una piccola lagena, che specialmente nei
Teleostei spesso contengono otoliti assai voluminosi, queste caratteristiche concrezioni di natura calcarea si
accrescono per opposizione superficiale in maniera discontinua, perchÚ il loro aumento di volume di norma si sospende nei mesi invernali.
Osservati in sezione presentano unĺalternanza di cerchi chiari e cerchi scuri attorno ad un nucleo centrale, i cerchi chiari essendosi deposti
in primavera-estate e quelli scuri estate-autunno (valutazione dell'etÓ).
Il labirinto e il sistema laterale hanno origine dalla
medesima origine embrionale, i rispettivi recettori sensoriali hanno delle strutture simili o identiche.
Infatti, nella macula delle creste
del labirinto e nei bottoni del sistema laterale, i recettori sensoriali sono fatti da uno o pi¨
fascetti di cellule piriformi, da dove si eleva
un ciuffo di flagelli non contrattili (uno per cellula)
incluso in un gel che le da la forma di cono alla cupola.
Medesima importanza pu˛ esser data
alle secrezioni mucose. Nellĺorecchio interno, la perilinfa Ŕ vischiosa, il muco della linea laterale e del labirinto hanno quasi la medesima
composizione (mucoproteine).
Funzioni del sistema laterale
I pesci apprezzano la corrente dell'acqua grazie alla linea laterale.
Un oggetto in movimento nell'acqua rompe l'equilibrio del mezzo liquido e
produce anche delle vibrazioni, la rottura dell'equilibrio causa delle correnti che esercitano delle deboli pressioni sui recettori sensoriali.
Le vibrazioni meccaniche agiscono nella medesima maniera ma le variazioni di pressione che essi creano sono ritmiche,
cosa che pu˛' esser
importante per gli stimoli deboli in quanto agiscono per sommatoria.
Nei canali sottocutanei il cambiamenti di pressione si trasmetteranno mediati dal muco.
Il senso dell'equilibrio
I pesci rispondono ai cambiamenti della posizione del corpo in rapporto alla direzione della forza di gravitÓ con dei riflessi statici e dei riflessi dinamici:
-
I riflessi statici
Consistono in delle posizioni compensatrici degli occhi, della testa, delle pinne, del tronco,
riferite a dei cambiamenti
d'orientamento del pesci in rapporto alla verticale.
-
I riflessi dinamici
Consistono in dei movimenti con effetto compensatorio, che sono provocati da accelerazioni angolari durante le rotazioni e da
accelerazioni lineari durante le traslazioni lineari. Qust'ultimi sono causati dall'azione subitanea dei muscoli effettori e differiscono
dai riflessi statici per la brevitÓ del loro periodo di latenza.
L'organo auditivo Ŕ collegato a quello dell'equilibrio ed innervato con l'ottavo nervo cranico, che per tale ragione Ŕ denominato nervo stato-acustico.
L'insieme dell'organo Ŕ collegato sul retro del cranio in una capsula cartilaginosa od ossea e costituisce, insieme alla capsula olfattiva e visiva,
una delle parti fondamentali del cranio dal punto di vista evolutivo.
Le afferenze dell'orecchio interno sono
strettamente correlate alla linea laterale per la sensibilitÓ a variazioni di pressione e
movimenti di correnti (orientamento tattile a distanza): si tratta di gruppi di cellule sensitive secondarie, meccanorecettrici, i neuromasti,
innervate dal 10░ paio di nervi cranici (nervo vago).
Sembra che i riflessi statici siano la risposta all'eccitamento alla
macula dell'utricolo e i riflessi dinamici all'eccitamento delle creste dei canali semicircolari.
Funzione degli organi a otoliti nel senso dell'equilibrio
Questi organi hanno in comune le concrezioni calcaree, che possono opprimere lo strato di muco che circonda i flagelli delle cellule
sensoriali.
L'organo auditivo Ŕ collegato a quello dell'equilibrio ed innervato con l'ottavo nervo cranico, che per tale ragione Ŕ denominato
nervo
stato-acustico.
L'insieme dell'organo Ŕ collegato sul retro del cranio in una capsula cartilaginosa od ossea e costituisce, insieme alla capsula olfattiva e
visiva, una delle parti fondamentali del cranio dal punto di vista evolutivo.
L'otolita esercita una duplice stimolazione:
- preme sempre seguendo la verticale, secondo la gravitÓ, le cupole sottostanti (funzione statica)
- a ogni movimento del pesce, lĺotolita si sposta sulla macula e in certi punti ne eccita le cupole secondo certe direzioni (funzione dinamica).
I movimenti sono cosi percepiti immediatamente dal pendulo a torsione dei canali semicircolari e dagli organi a otoliti del labirinto.
Gli
organi a otoliti sono sensibili anche a vibrazioni sonore (funzione acustica)
U. Fazzini
2003-2004
(Testo integrato con AA)
Altro
Il sistema acustico-laterale
Lĺudito
La
velocitÓ di propagazione del suono nell'acqua Ŕ circa quattro volte superiore a quello nell'aria:
infatti il suono nell'acqua compie 1500 metri al secondo.
Un pesce pu˛ determinare la distanza da cui proviene il suono e la
direzione in cui questo si sposta e ci˛ nel
mezzo acquatico Ŕ particolarmente importante considerando la
scarsa funzionalitÓ della visione a distanza.
Produzione di suoni
Specie della famiglia Pomadasyidae ad esempio, producono un suono acuto sfregando una
placca di denti faringei contro alcune placche ventrali denticolate pi¨ piccole.
Nei Balistidae
suoni particolari vengono prodotti sfregando le spine anteriori fuse della pinna dorsale.
Alcune
specie del genere Mycteroperca battono lĺopercolo contro il corpo per produrre piccoli tonfi.
Negli Sciaenidae, Triglidae e Gadidae la compressione del gas presente nella vescica
natatoria consente loro di produrre suoni brevi e di bassa intensitÓ.
In alcuni Gadidi come Melanogrammus
aeglefinus sequenze di suoni stereotipate vengono emessi dal maschio nel corso del
comportamento nuziale.
Otoliti
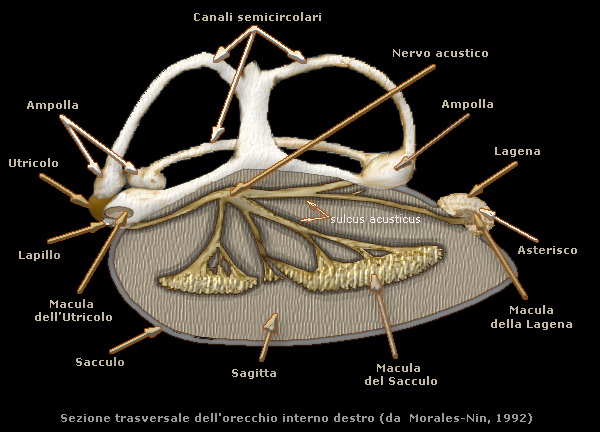
Con
il termine otolite si indica, generalmente la sagitta che Ŕ il pi¨ grande degli ossicini e
viene deposto nel sacculo; nellĺutricolo avviene la formazione del
lapillo, mentre nella lagena si forma
lĺasterisco.
Le dimensioni della sagitta possono superare anche il centimetro, mentre asterisco e
lapillo sono pi¨ piccoli misurando soltanto pochi millimetri con il primo pi¨ grande del secondo.
La sagitta possiede un corpo ellittico lateralmente compresso e
concavo distalmente con lĺasse lungo orientato in direzione antero-posteriore. Il lato convesso
o prossimale, orientato verso lĺasse centrale, Ŕ diviso in due aree da un solco profondo, il
sulcus
acusticus, sul quale si inseriscono le fibre nervose provenienti dalle pareti del sacculo.
Dal punto di vista chimico, gli otoliti sono composti da
cristalli di carbonato di calcio (CaCO3), sotto forma di aragonite, e da una matrice organica, a sua volta
costituita in gran parte da una proteina ricca di aspartato e glutammato chiamata otolina.
La macula dellĺutricolo e, in minor grado, quelle del sacculo e
della lagena registrano, attraverso lĺinclinazione dellĺotolite e dei prolungamenti sensitivi in
esso inclusi, lĺinclinazione della testa e
lĺaccelerazione lineare. La sagitta e lĺasterisco sembrano
essere principalmente coinvolti nella percezione dei suoni.
Le onde sonore fanno vibrare queste
strutture e ancor di pi¨ lĺepitelio sensoriale delle rispettive macule provocando la trasmissione di
stimoli nervosi al centro acustico del cervello.
Negli Elasmobranchi gli otoliti consistono di una matrice
gelatinosa contenente numerosi piccoli cristalli di carbonato di calcio.
A.A.
Altro
L'organo stato-acustico bilaterale o labirinto, situato dorsalmente e lateralmente al midollo allungato, Ŕ completamente
incorporato nell'osso cranico.
Ogni metÓ del labirinto Ŕ costituita da tre canali semicircolari (orientati nei piani orizzontale, laterale e longitudinale) e da tre
camere otolitiche (utricolo, sacculo e lagena).
I canali semicircolari e l'utricolo costituiscono la parte superiore dorsale, responsabile del
mantenimento dell'equilibrio spaziale e del bilanciamento, mentre il complesso sacculo-lagena costituisce la parte inferiore ventrale,
responsabile della rilevazione dei suoni.
I canali semicircolari sono rivestiti da epitelio squamoso semplice e sono riempiti di endolinfa fluida che si muove liberamente all'interno
del lumen del canale. Alla loro giunzione con l'utricolo, i canali semicircolari si allargano in ampolline (camere di rilevamento
dell'inerzia del fluido).
All'interno di ciascuna ampolla si trova una cresta
ampollare rivestita da cellule epiteliali sensoriali globose e
che sostiene una macula rialzata di cellule ciliate sensoriali avvolte da una cupola gelatinosa.
La cupula si proietta nel percorso del fluido all'interno del canale.
Il movimento del pesce induce un ritardo
inerziale nell'endolinfa. Il risultato Ŕ un movimento apparente del fluido all'interno del canale nella direzione opposta al movimento
effettivo del pesce. Lo spostamento della cupola nella direzione del movimento del fluido provoca una deflessione dei peli sensoriali.
Le
cellule sensoriali comunicano le informazioni sul movimento al cervello attraverso la porzione anteriore del VII nervo cranico.
Come i canali semicircolari, le camere otolitiche sono rivestite di epitelio squamoso e riempite di endolinfa. Le macule sono appiattite
sulla superficie ventrale delle camere, sopra le quali (e a contatto con i peli delle cellule sensoriali) sono sospesi gli otoliti. Le onde
sonore in campo vicino, che si propagano sotto forma di vibrazioni, provocano lo spostamento delle particelle d'acqua.
I pesci sottoposti a
queste vibrazioni si muovono di concerto con le particelle d'acqua circostanti. La densitÓ comparativa degli otoliti rispetto all'endolinfa
circostante (circa 3:1) fa sý che essi rimangano fermi rispetto al movimento laterale del pesce. La conseguente deflessione dei peli induce
le cellule sensoriali a inviare informazioni uditive al cervello attraverso la porzione posteriore del VII nervo cranico.
Le onde sonore in campo lontano si propagano nell'acqua come onde di pressione. PoichÚ non generano il movimento delle particelle, queste
onde non producono il movimento avanti e indietro necessario per il rilevamento dell'otolite.
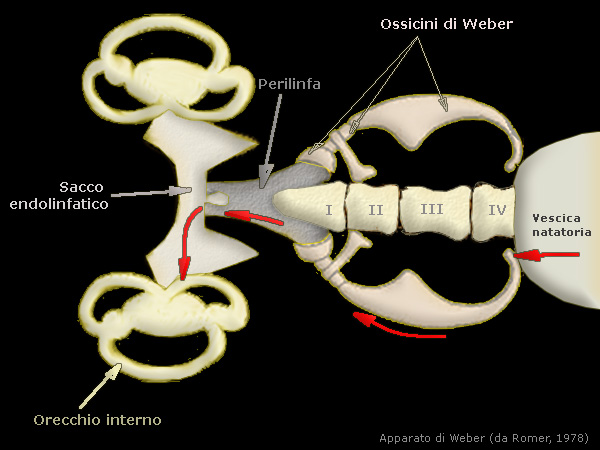
L'amplificazione delle onde di campo lontano,
tuttavia, avviene all'interno delle camere a bassa densitÓ della vescica gassosa, provocando vibrazioni della parete della vescica. Queste
vibrazioni vengono tradotte attraverso l'apparato weberiano agli organi stato-acustici.
I processi neurali delle quattro vertebre pi¨ anteriori
si staccano e si sviluppano indipendentemente dai loro centri per formare l'apparato weberiano.
Queste ossa e i legamenti associati
forniscono un collegamento rigido tra la camera anteriore della vescica gassosa e la parte inferiore (complesso sacculo-lagena) dell'organo
stato-acustico. La trasduzione delle onde di pressione in vibrazioni nella vescica gassosa e la traduzione di queste vibrazioni alla parte inferiore aumenta notevolmente la sensibilitÓ uditiva.
A.A.
|


















