 |
Pesci -
Anatomia - Organi Respiratori
|
|
Branchie
Struttura e funzione |
La grande maggioranza delle specie di
pesci utilizza le branchie per la respirazione. Poche specie sono in
grado di estrarre ossigeno dall’aria, e possono impiegare a tale
scopo le branchie, la vescica natatoria ed altri organi respiratori
accessori inclusa la pelle.
Le branchie sono composte essenzialmente
da un complesso sistema di vasi sanguigni, contornati da un epitelio
che fornisce una sottile barriera tra il sangue e l’ambiente
acquatico.
L’intero flusso sanguigno entra nel sistema vascolare
branchiale, si ossigena, e ritorna nel sistema circolatorio
attraverso l’aorta dorsale. Le caratteristiche morfo-funzionali
delle branchie ne fanno degli organi molto efficienti non solo per
la respirazione ma anche per altre funzioni fisiologiche.
L’apparato
branchiale č posto ai lati della faringe.
Nei selaci la faringe su
ciascun lato presenta una serie di fessure, quelle del primo paio,
sono di piccole dimensioni e formano lo spiracolo, seguono altre 5
fessure branchiali di maggiori dimensioni. Le branchie sono poste
sulle pareti delle fessure. L’acqua fluisce dalla bocca alla faringe
e fuoriesce dalle fessure branchiali, durante questo passaggio
avvengono gli scambi gassosi con il torrente sanguigno.
Nei teleostei le branchie, poste in una camera branchiale, ai due lati
della faringe, sono protette da un opercolo, formato da lamine
ossee.
Nella respirazione gli opercoli si chiudono e la cavitŕ
boccale si dilata, aspirando acqua nella bocca. Contemporaneamente,
le camere branchiali si dilatano in modo da far fluire l’acqua sulle
branchie facendola fuoriuscire dagli opercoli.
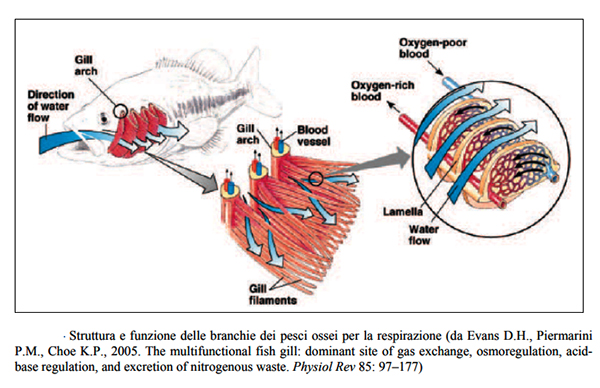
Ciascuna branchia č
sostenuta da un arco osseo branchiale che porta quattro olobranchie.
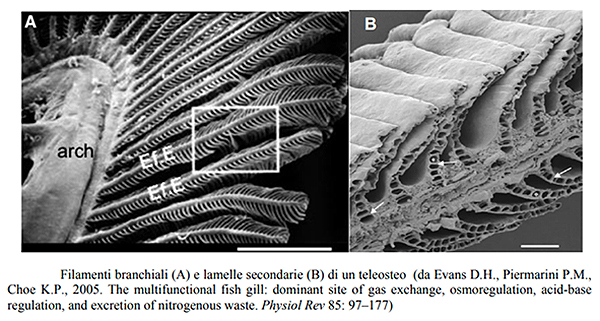
Queste sono costituite da filamenti sottili detti filamenti
branchiali sui quali sono allineate in modo ordinato delle
formazioni trasversali a forma di semiluna dette lamelle secondarie.
Ciascun filamento č rifornito di sangue a basso tenore
di ossigeno attraverso un’arteria afferente, che si estende lungo
tutto il filamento. Il sangue da questa arteria fluisce in rete di
capillari presenti all’interno delle lamelle secondarie e una volta
ossigenato, dal flusso d’acqua che scorre negli spazi tra le
lamelle, ritorna in circolo attraverso un’arteria afferente che
decorre lungo il filamento branchiale, parallelamente all’arteria
efferente. Le lamelle aumentano enormemente l’area superficiale
delle branchie incrementandone l’efficienza come organi respiratori.
La capacitŕ delle branchie di estrarre ossigeno
dall’acqua č inoltre aumentata dal fatto che il sangue nelle lamelle
scorre controcorrente rispetto al flusso d’acqua. Il sangue che
entra nei filamenti branchiali e quindi nelle lamelle ha una minore
pressione parziale di ossigeno rispetto all’acqua. Questa ultima
scorrendo controcorrente, cede gradualmente O2 al sangue
ossigenandolo. Nel momento in cui il sangue lascia la lamella
secondaria la sua pressione parziale di O2 ha quasi raggiunto quella
dell’acqua.
L’epitelio branchiale non svolge
solamente un ruolo per gli scambi gassosi ma ha una funzione
fondamentale per l’osmoregolazione e l’escrezione di cataboliti
azotati.
I pesci che vivono in mare sono iposmotici rispetto
all’acqua marina, la loro concentrazione osmotica č infatti 1/3
rispetto a quella dell’acqua di mare e tendono quindi a
disidratarsi.
Teleostei e elasmobranchi hanno evoluto strategie
differenti per bilanciare la perdita di acqua e l’acquisizione per
diffusione di NaCl attraverso le branchie.
I teleostei evitano la
disidratazione ingerendo acqua marina, che viene poi assorbita
dall’epitelio intestinale, determinando in questo modo un aumento
del carico di sali che va a sommarsi a quello che penetra per
diffusione dalle branchie.
L’eccesso di sali assorbito č
successivamente eliminato per trasporto attivo di Na+, Cl
- e forse K+ attraverso l’epitelio delle branchie (cellule cloruro) e per
escrezione di sali bivalenti operata dal rene.
Negli elasmobranchi l’ingestione di acqua non č necessaria perché il
plasma č reso iperosmotico rispetto all’acqua marina da una elevata
ritenzione di urea. Ciononostante, squali e razze, sono costretti ad
espellere l’eccesso di NaCl che entra attraverso le branchie per
mezzo di una ghiandola rettale che deriva dall’intestino posteriore.
Nei pesci, le branchie sono inoltre il principale sito di escrezione
di cataboliti azotati, ammoniaca e urea in primo luogo. Oltre l’80%
di queste sostanze vengono escrete attraverso le branchie.
Quasi
tutti gli actinopterigi e gli agnati, espellono l’azoto in eccesso
come ammoniaca e sono detti ammoniotelici. L’ammoniaca č, infatti,
altamente tossica e non puň essere “conservata” nei tessuti. Il
principale effetto dell’ammoniaca č quello di sbilanciare i
gradienti elettrochimici a livello del sistema nervoso centrale, per
l’abilitŕ della sua forma ionica, lo ione ammonio NH4 + di
sostituirsi allo ione potassio K+ nei trasportatori ionici.
Gli elasmobranchi, i celacanti e alcuni teleostei producono urea e sono
detti urotelici. Questo adattamento sembra
essersi evoluto sia come un meccanismo di osmoregolazione in acqua
di mare che per la detossificazione dell’ammoniaca.


















