 |
Pesci - Anatomia - Organi Respiratori
|
|
Branchie
Escrezione di ammoniaca |
Il catabolismo degli aminoacidi e delle basi azotate genera, per
la maggior parte, rifiuti essenzialmente rappresentati
dall'ammoniaca (NH3). che č molto tossica e non puň essere
accumulata nei tressuti. I principali effetti dell'ammoniaca
sono quelli di sbilanciare i gradienti elettrochimici a livello del sistema nervoso centrale
e di sostituirsi (come ione ammonio) allo ione potassio nei trasportatori ionici.
Nei pesci, le branchie sono il principale sito di escrezione di questi rifiuti.
Quasi tutti gli actinopterigi e gli agnati, espellono l'azoto in eccesso come ammoniaca e sono detti
ammoniotelici.
Gli elasmobranchi, i celacanti e alcuni teleostei producono urea
e sono detti urotelici. Questo adattamento, oltre ad essere
osmoregolatore, si č evoluto sia come detossificatore dell'ammoniaca.
I teleostei sono organismi ammoniotelici. Infatti, espellono circa 95%
dei loro rifiuti di azoto sotto forma di ammoniaca. Il rimanente 5% dei
rifiuti viene trasformato in acido urico e urea ed č eliminato dal
sistema escretore renale.
L'ammoniaca si trova sotto forma di ioni ammonio NH4+ a pH
fisiologico e viene escreta quasi tutta attraverso le branchie,
piů o meno alla stessa velocitŕ della sua produzione, E' il
terzo gas respiratorio ed č deputato a svolgere un ruolo nel controllo
della respirazione.
Poiché gli ioni di ammonio difficilmente
possono essere escreti per semplice
diffusione, il meccanismo del loro smaltimento richiederebbe almeno un trasportatore.
In studi passati si affermava che le sostanze di piccola
massa molecolare, come NH3,
CO2 , urea e acqua, fossero considerate relativamente
permeabili, quindi non avrebbero necessitato di alcun trasportatore o canale
specializzato, ma semplicemente sarebbero scivolate oltre o dissolte
nei lipidi di membrana nel doppio strato lungo il gradiente di
diffusione.
La
diffusione non ionica di NH3
veniva suggerita da un efficace complesso di scambio Na+ /NH4+
(antiporto branchiale),
che in effetti esiste sulla superficie apicale dell'epitelio
branchiale e che probabilmente fa ricorso a diversi trasportatori di membrana, invece che da
un singolo scambiatore Na+ /NH4+
Al contrario, le molecole ioniche (ad esempio NH4+, Na+, Cl– richiedevano trasportatori/canali proteici attivi (ad esempio
Na/K+-ATPasi)
o passivi (ad esempio scambio HCO3- /Cl- ,
canale Na+).
Randall e Tsui (2002) e Wood (1993) avevano ipotizzato un meccanismo per il trasporto attivo dello ione ammonio
attraverso l’epitelio delle branchie di pesci teleostei con il coinvolgimento di
due ATPasi
di membrana, la Na,K-ATPasi basolaterale e una
Na,H-ATPasi
apicale, che avrebbero trasportato lo ione ammonio rispettivamente in
sostituzione di K+ e di H+.
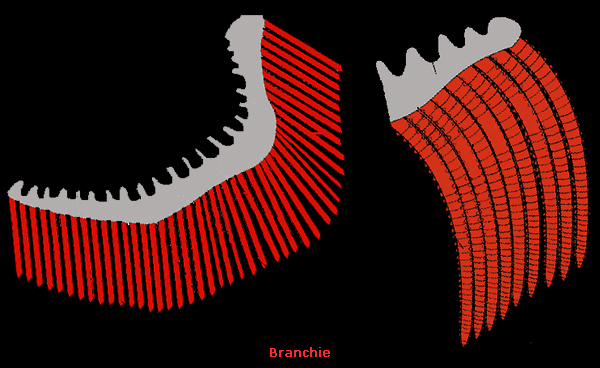
In questo meccanismo le cellule del cloruro
avrebbero messo in opera un
assorbimento netto (spostamento dall’esterno all’interno
dell’organismo) di Na+
e Cl-.
L’assorbimento di NaCl
sarebbe accoppiato all’estrusione di scorie azotate (coppia NH4+
e NH3)
e alla regolazione dell’equilibrio acido-base.
Le cellule del
cloruro, cioč, svolgerebbero, negli Osteitti d’acqua dolce, le funzioni
che nell’uomo sono distribuite tra reni, canale alimentare e
polmoni.
Sulla membrana basolaterale sarebbe localizzata la Na+
/K+ -ATPasi che manterrebbe un gradiente elettrochimico favorevole
all’ingresso in cellula di Na+.
In acqua marina cambiarebbe il verso degli spostamenti,
perché differenti sono i trasportatori della membrana apicale e
di quella basolaterale che verrebbero impiegati.
Questi meccanismi sarebbero
sovrapposti ad un sostanziale movimento verso l'esterno di NH3
per semplice diffusione, che sarebbe dipesa dall'intrappolamento
acido nell'acqua dello strato limite da parte di ioni H+ creati
dall'idratazione catalizzata o non catalizzata della CO2 metabolica
espirata.
Tuttavia, dal 2000, il paradigma č cambiato con la
scoperta che, nei mammiferi, le glicoproteine Rh
possono fungere da canali selettivi che facilitano notevolmente
la diffusione dell'ammoniaca attraverso le membrane cellulari.
Tale meccanismo č stato scoperto anche nelle
branchie dei granchi e in alcuni telostei.
La loro
importanza nell'escrezione di ammoniaca č stata rafforzata
dalla scoperta che i livelli di espressione di mRNA delle
proteine Rh nelle branchie erano sovraregolati,
nella trota,
dal carico di ammoniaca esterno o interno e che, nel pesce
zebra,
l'abbattimento molecolare dell'espressione della proteina Rh
riduceva gravemente l'escrezione di ammoniaca nelle larve.
La funzione di trasporto
dell'ammoniaca delle proteine Rh della trota č stata
definitivamente dimostrata da studi di espressione degli
ovociti in vitro.
Le proteine Rh svolgono un ruolo chiave nei modelli
attuali dell'escrezione branchiale di ammoniaca dei pesci teleostei.
Mentre una parte
di ammoniaca puň passare per semplice diffusione attraverso
le membrane cellulari delle lipoproteine, una parte
significativa puň anche passare per diffusione facilitata
attraverso i canali delle proteine Rh, e si pensa che
questa frazione aumenti quando l'espressione della proteina
Rh č indotta dal carico di ammoniaca.
In effetti, viene
proposto che il trasporto attivo verso l'esterno di
ammoniaca spieghi
l'aumentata escrezione di ammoniaca dopo l'alimentazione,
l'esercizio fisico o l'infusione di ammoniaca, nonché il
ripristino dell'escrezione di ammoniaca quando i normali
gradienti pNH3 e NH4+ elettrochimici diretti verso l'esterno
vengono invertiti da elevati livelli di ammoniaca esterna.
Basato sulla partecipazione attiva di un metabolone mediato da geni delle glicoproteine Rhesus
(Rh) e collegato
all'assorbimento di Na+ e all'escrezione acida, un nuovo modello per l'escrezione di ammoniaca nei
pesci d'acqua dolce apre la strada a nuovi meccanismi
escretori.
In questo modello,
- Rhag facilita il flusso
di NH3 fuori
dall'eritrocita,
- Rhbg la sposta attraverso la membrana basolaterale dell'ionocita branchiale,
- un complesso di
scambio Na+ /NH4>+ apicale, costituito da diversi trasportatori di membrana (Rhcg,
H+ -ATPasi di tipo V, scambiatore Na+ /H+
NHE-2 e/o NHE-3, canale Na+) fornisce un meccanismo di intrappolamento acido per l'escrezione apicale.
- l'anidrasi carbonica intracellulare (CA-2) e il cotrasportatore basolaterale Na+ /HCO3- (NBC-1)
e la Na+ /K+ -ATPasi svolgono ruoli indiretti.
Su queste basi, in futuro, si apriranno vaste indagini
sulle proteine Rh nei pesci, che vedranno il loro coinvolgimento nel
meccanismo di escrezione di ammoniaca attraverso le branchie nei
pesci d'acqua marina, la loro possibile importanza
nell'escrezione di ammoniaca attraverso la pelle, il loro
potenziale duplice ruolo come trasportatori di CO2 ,
le loro risposte all'alimentazione e i loro ruoli nelle prime
fasi della vita prima del pieno sviluppo delle branchie.
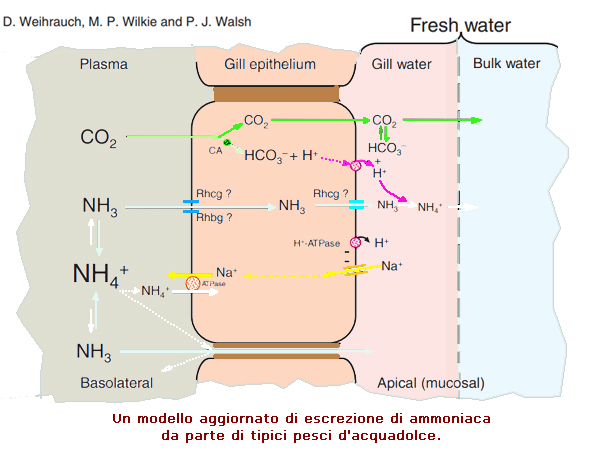
Quando la CO2 viene escreta attraverso le branchie, viene
idratata nell'acqua delle branchie (strati limite non agitati)
per generare H+ e HCO3–.
L'H+ risultante generato dall'idratazione della CO2 e
probabilmente dall'attivitŕ apicale dell'ATPasi H+, intrappola
l'NH3 come NH4+, poiché diffonde passivamente nell'acqua delle
branchie, mantenendo il gradiente PNH3 transcellulare.
Le prove genomiche e fisiologiche emergenti suggeriscono che il
trasporto di ammoniaca attraverso la membrana plasmatica delle
cellule branchiali dipende dalla presenza di glicoproteine Rhesus.
Sulla base di questa evidenza si ipotizza che le glicoproteine
Rhcg o Rhbg sulla membrana basolaterale agiscano come condotto
per il trasporto di NH3 (le glicoproteine Rh sono anche
permeabili a NH4+) nel citosol delle cellule branchiali, seguito
dalla diffusione di NH3 verso l'esterno tramite le glicoproteine
Rhcg apicali.
Rimane anche la possibilitŕ che una unica NH4+- ATPasi Na+
dipendente -, ancora non caratterizzata, contribuisca anche al
trasporto di ammoniaca basolaterale (Salama et al., 1999).
A causa della presenza di profonde giunzioni strette tra cellule
adiacenti nella branchia d'acqua dolce, sembra improbabile che
vi sia una diffusione paracellulare apprezzabile di NH4+ nei
pesci d'acqua dolce.
Testo adattato
Fonti
-
Sandrine Heusser -
Licence de sciences de la vie
-
Faculté des sciences et techniques Université Jean Monnet de
Saint-Étienne -
Département de biologie-biochimie-
2017
-
A new paradigm for ammonia excretion
in aquatic animals: role of Rhesus (Rh) glycoproteins-
Patricia A. Wright1, and Chris M. Wood - pdf
-
Ammonia and urea transporters in gills
of fish and aquatic crustaceans
Dirk Weihrauch, Michael P. Wilkie and Patrick J. Walsh
- pdf
-
Ammonia production, excretion,
toxicity, and defense in fish: a review -
Yuen K. Ip, and Shit F. Chew - pdf
-
Appunti di Ecofisiologia animale -
Giuseppe Cassano - pdf
-
Is ammonia excretion affected by gill
ventilation in the rainbow trout Oncorhynchus mykiss?
Junho Eom, Sandra Fehsenfeld, Chris M. Wood - pdf
-
Organismi acquatici e ambiente:
meccanismi biochimici di interazione, risposta e adattamento
(Tesi)
Patrizia Bandiera - pdf
-
Rhesus glycoprotein gene expression in
the mangrove killifish Kryptolebias marmoratus exposed to
elevated environmental ammonia levels and air
C.Y. C.Hung, K.N.T.Tsui , J.M.Wilson , C.M.Nawata, C.M.Wood
and P.A.Wrigh - pdf


















