|
 |
Pesci - Apparato
muscolare |
|
T.M. - Organizzazione muscolatura
Miomeri e Miosetti |
Organizzazione della muscolatura
La muscolatura somatica e del tronco e della coda consta;come
già detto, di una serie di miomeri
e miosetti.
I miomeri sono
perpendicolari al corpo ma hanno una struttura complicata.
Per una migliore
efficienza, in quanto devono sviluppare una forza ottimale e veloce durante la
contrazione, l’inserzione della fibra muscolare può assumere angoli diversi
rispetto all’asse del corpo, quest’angolazione dipende dal tipo di pesce e dal
suo modo di nuotare veloce, lento o continuo
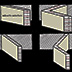
Una semplice inclinazione rispetto all’asse e in direzione del
piano mediano non sarebbe sufficiente, poiché i punti d’attacco dei miomeri nel
tegumento e nello scheletro assile possono esser spostati durante la
contrazione.
Ogni miomero appare, quindi, ripiegato come la lettera
W ed
è disposto orizzontalmente. I miomeri s’incurvano non soltanto sul piano
sagittale, ma anche in quello trasversale, quindi formano dei coni con i vertici
rivolti in senso cefalico e caudale (coni dorsali e coni ventrali).
Questa
disposizione permette la trasmissione delle trazioni, per mezzo dei miosetti, a
tutta la colonna vertebrale.
Quest’organizzazione, tipica nella regione
caudale, si mantiene nella regione dorsale del tronco mentre nella parte
ventrale si modifica in rapporto alla formazione della cavità viscerale.
Questa è
delimitata da uno strato superficiale e uno profondo e si differenziano i
muscoli:
- obliquo esterno
- obliquo interno
- retto laterale
- retto mediale.
Nella regione branchiale la muscolatura si differenzia e,
spostandosi in senso craniale, forma la muscolatura ipobrachiale ed
epibrachiale .
Durante la contrazione, il punto mediale o laterale
d’inserzione rimangono in un piano paratrasversale e solo l’angolo posto
all’interno diventa meno acuto, il muscolo può così contrarsi senza scuotere la
parte esterna del corpo.
In pesci come i selaci e lo storione, i miomeri
assomigliano ad una W inclinata, il cui margine superiore è orientato
verso la testa.
Gli strati sottostanti hanno una struttura alquanto complessa.
Nei Teleostei il numero dei miomeri è normalmente identico al numero delle
vertebre, benché è difficile contarne il numero vicino alla base delle pinne
caudali poiché le vertebre sono spesso modificate in quella regione.
Nei Ciprinidi sono sempre due miomeri in più del numero delle vertebre in quanto due
vertebre sono modificate per contribuire alla formazione della regione
occipitale delle ossa della testa.
Negli Squali possono esserci due miomeri per
ogni vertebra.
L’intera muscolatura somatica origina dal cranio e dal cinto
pettorale.
La porzione epiassiale origina dalla volta del neurocranio e dalla
parte dorsale del cinto pettorale.
L’ipoassiale solo dalla parte ventrale del
cinto dato che lo scheletro branchiale non è in continuità con il cranio.
La
base della pinna caudale, i processi spinosi e le coste sono i punti
d’inserzione di tali muscoli.
La forza di contrazione dei miomeri anteriori è
trasmessa ai sempre più piccoli miomeri verso la fine del corpo, dove dopo aver
agito sui tendini va verso le pinne caudali.
Il diametro delle fibre muscolari
può avere una gran variabilità nei vari muscoli.
Nelle varie specie, il diametro
delle fibre slow varia tra i 69 e 75 mm, mentre quello
delle fast tra i 35 e 45 mm; la lunghezza del sarcomero (tra due strie Z)
dipende dal tipo di fibra nelle slow ha una media di 1,82 mm mentre nelle
fast
di 2,04 mm.
La vascolarizzazione varia considerevolmente dipendendo dal tipo di
fibra (fast o slow) dalla porzione muscolare e dal tipo di pesce.
Per esempio
nella perca fluviale nella muscolatura della testa vi sono 167 fibre (diametro
27-74 mm) e 76 capillari per mm2 nella regione della coda 281 fibre
(diametro 24-63 mm) e 139 capillari sempre per mm2, in quella del
tronco 1084 fibre (diametro 13-34 mm) e 1257 capillari, nella coda 3022 fibre
(diametro 11 - 23 mm) e 4800 capillari.
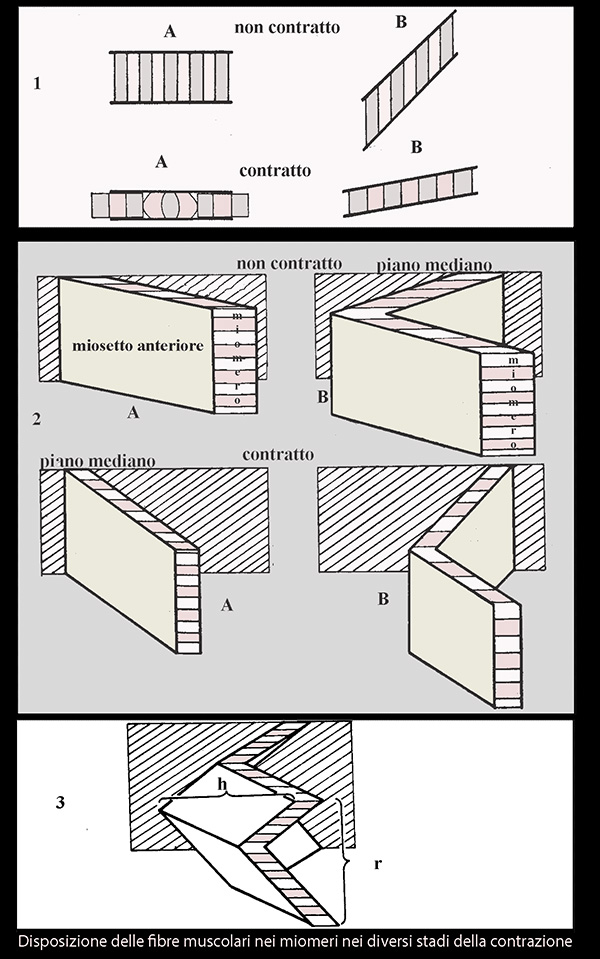
Un sistema con
diversa organizzazione di muscoli muove la lama delle pinne mediali caudali,
anali e dorsali nonché delle pinne pari quali le pettorali e le pelviche, mentre
un sistema addizionale muscolare aprendo i raggi controlla la forma dalla lamina
della pinna.
I muscoli principali nelle pinne
pettorali sono l’adduttore superficiale e profondo e il blocco degli
abduttori che s’inseriscono con base a forma di
T nei raggi delle pinne
pari. Il primo raggio, che può esser soggetto alla maggiore resistenza del
flusso, ha anche muscoli estensori e flessori.
Similmente i muscoli
erettore e flessore delle pinne mediane si legano ai raggi fino ai pterigiofores. Vari muscoli poi controllano la
distensione delle pinne come ad esempio i muscoli inclinatori delle pinne
dorsali e ed anali e i muscoli erettori delle pinne pettorali.
Nell’evoluzione delle pinne caudali degli actinopterigi c'è la
tendenza all’aumento della complessità dei muscoli per il controllo
dell’orientamento e forma della pinna.
Nei selaci e
nei primitivi actinopterigi i piegamenti della pinna caudale sono ottenuti
mediante la contrazione dei muscoli miotomali con i muscoli superficiali
laterali che s’inseriscono direttamente nei raggi della pinna caudale.
Movimenti
indipendenti dai muscoli miotomali dei raggi della pinna caudale li troviamo nei
Ginglimodi, principalmente per i raggi ipoassiali per lo sviluppo di un debole
muscolo longitudinale e uno sviluppato muscolo flessore ventrale.
Muscoli antagonisti interadiali e sopracrarinali originatesi negli
Halecostomi provvedono ad aumentare il controllo d’apertura delle pinne.
I teleostei hanno muscoli epiassiali ed ipoassiali
interradiali e carinali. Il muscolo longitudinale ipocordale è inoltre ben
sviluppato e probabilmente è critico nell’irrigidire e stabilizzare la porzione
epiassiale della pinna caudale durante le partenze rapide.
I fasci muscolari delle pinne degli actiniperigi tendono ad esser
piccole e fusiformi o moderatamente appiattite, mentre quelle dei batoidiformi
tendono ad esser più grandi ed organizzate in lamine.
Le fibre superficiali rosse nelle pinne pettorali sono orientate
parallele ed inserite all’incirca alla metà dei ceratorichi.
Le fibre bianche sono disposte obliquamente ai ceratorichi con una
terminazione nastriforme con una probabile conseguenza di aumentare la forza di
spinta.
L’anatomia e l’organizzazione delle fibre dei miotomi è complessa comparata a quella riscontrabile nei muscoli delle pinne
mediane e pari.
Le fibre bianche hanno due tipi di
traiettorie
- Una è associata ai tendini o equivalenti, e si continua nei
miosetti dei selaci e in minor misura in alcuni actinopterigi (acipenser,
anguilla, salmonidi) e nella regione caudale della maggior parte dei
teleostei.
- La seconda con le traiettorie dei fasci muscolari ad
andamento ad elica è presente nei miomeri anteriori dei teleostei dove sono
assenti strutture tendinee.
Tutte le fibre rosse sono
approssimativamente equidistanti
dall’asse mediano del corpo e ad esso parallele. Tutti i sarcomeri si accorciano
in eguale misura nel range di lunghezza con generazione di forza > 96% della
massima potenzialità.
Diversamente le fibre bianche variano in distanza dal
piano mediale e sottendono angoli da 10° a quasi 40 ° dall’asse del
corpo. Quest’organizzazione assicura che tutte le fibre bianche si contraggano
con proporzione similare della loro lunghezza, sviluppando forze
superiori all'85% del
massimo, mentre lavorano nella porzione della curva dove potenza ed efficienza
sono alti.
Le traiettorie ad elica consentono alti
ritmi di curvatura comparati a quelli determinati dall’organizzazione delle
fibre rosse. Ciò ha importante conseguenza per adeguare il rapporto delle fibre,
dove questo cambia nella posizione del corpo per un determinato cambiamento
nella lunghezza del sarcomero.
L’adeguamento della
fibre bianche è quattro volte maggiore delle fibre rosse, il tutto combinato con
la Vmax più alta, risulta che le fibre bianche producano movimenti di un
ordine di grandezza maggiore rispetto alle fibre rosse.
I piegamenti dei miomeri avvenendo non solo in singolo piano ma in un complesso di strutture
tridimensionali, permettono alle fibre muscolari di scorrere parallelamente al
piano orizzontale e mediano. Per mettere in grado di avere una contrazione
uniforme, queste sono della medesima lunghezza (l’estensione della contrazione è
coordinata dal sistema nervoso).
Le fibre devono altresì attaccarsi ai miosetti
con la medesima angolazione, o con angoli che sono messi in relazione gli uni
agli altri in maniera definita. Perciò le fibre muscolari scorrono obliquamente
in rapporto al piano principale del corpo. Il grado d’inclinazione è usualmente
di 30°, sezioni parallele al piano mediale od orrizzontale, perciò, raramente
contengono singole fibre per la loro intera lunghezza. Questo accade solo nel
muscolo superficiale del tronco, negli altri muscoli (muscolatura bianca)
comprende solo porzioni di fibre disposte in varie direzioni, le fibre muscolari
bianche presentano, infatti, due modelli di disposizione:
- La prima è caratteristica degli Elasmobranchi, Dipnoi,
Condroitti ed Oleostei nonché in alcuni Teleleostei come l’anguilla e i
salmoni (tipo Elasmobranchi)
- La seconda tipica della maggior parte dei Teleostei ma solo
per la muscolatura epiassiale e ipoassiale e nella sua porzione anteriore
della muscolatura della coda.
Nel tipo Elasmobranchi le traiettorie sono di due tipi:
- dai coni muscolari orientati in senso cranio caudale
s’inseriscono nelle aponeurosi
- sono orientate dalle aponeurosi del cono alla colonna
vertebrale o al setto mediano.
Nell’ultimo caso ciascuna traiettoria incrocia pochi miomeri,
circa otto nella parte anteriore del corpo e cinque nella parte terminale della
coda.
Nel tipo Teleostei le traiettorie vanno quasi come i segmenti
di una spirale circolare, questa forma un fascio di fibre coassiali può esser
comparata a quelli di una corda.
Quattro fasci sono disposti per ogni lato del corpo:
- Uno sottile verso il margine dorsale del corpo
- un’altro sottile verso la parte ventrale (fasci sussidiari)
- gli altri due, più larghi sono collocati tra i due
precedenti lungo il miosetto orizzontale.
I fasci ipoassiali sono più corti di quelli epiassiali, poiché
la cavità del corpo è coperta da muscoli obliqui che hanno uno sviluppo
indipendente dai modelli di fibra.
Ogni traiettoria consta di un segmento ad
elica circolare che compie un mezzo giro; il segmento inizia nel setto mediano
e, con andamento caudale, gira verso la parete del corpo e da qui ritorna verso
il setto mediano. L’angolo di inclinazione dell’elica del fascio principale e di
30°-45°, mentre quella dei fasci sussidiari è di circa 35°.
La lunghezza del
segmento ad elica con un dato angolo di inclinazione è circa quattro volte il
raggio dell’elica. La direzione di rotazione di un segmento ad elica è alternata
con i differenti fasci, così che, nello stesso tempo fasci principali scorrono
in direzioni opposte.
Le relative traiettorie iniziano vicino al miosetto
orizzontale: il fascio dorsale scorre dorsalmente, il ventrale
ventrocaudalmente.
Ciascun fascio sussidiario ruota sempre in modo contrario al
contiguo fascio principale, un fascio ha quindi una rotazione opposta a quello
omologo del lato controlaterale.
La direzione delle traiettorie risulta
dall’orientamento dei miomeri e delle proprietà fisiologiche delle fibre
muscolari. Ogni tipo di fibra ha una velocità di contrazione ed intensità
ottimali; il grado con cui si può contrarre con massima velocità ha un limite
oltre il quale la contrazione è meno intensa. Maggior forza è sviluppata durante
le contrazioni più rapide.
L’ordinamento delle fibre rappresenta un compromesso
tra la velocità e la forza ottimale, questo varia nelle differenti specie di
pesci secondo le loro habitat e modo di vita.
Il modello delle fibre tipo Teleostei è più favorevole ai rapidi movimenti, mentre il modello tipo
Elasmobranchi permette un più ampio grado di curvatura e flessibilità. Per
questo motivo la tipologia Elasmobranchi è presente solo nel peduncolo dei
Teleostei; questo, infatti, per considerazioni idrodinamiche, è sottile, ma allo
stesso tempo ha un'ampia flessibilità di movimento per creare la necessaria
forza propulsiva. L’ampiezza non potrebbe esser sufficiente con la muscolatura
tipo Teleostei, la frequenza sarebbe maggiore, ma il peduncolo dovrebbe avere
una più larga sezione per fornire spazio alla massa muscolare.
U. Fazzini
Università di Udine
2003-200
|


















