|
 |
Pesci -
Anatomia -
Organi
di Senso
|
|
Il Sistema Laterale |
Con il sistema acustico, quello laterale costituisce un insieme anatomico e
funzionale; ma mentre il primo č comune a tutti i vertebrati il sistema laterale
si limita ai vertebrati inferiori: agnati, pesci, anfibi (larve).
Puň esser
considerato come una specializzazione del sistema cutaneo generale, con le sue
strutture periferiche, per le sue fibre associate in nervi laterali e per i suoi
centri mielencefalici.
Il modo d’organizzazione dei recettori periferici
costituisce la caratteristica morfologica del sistema laterale; infatti, i neuromasti non
si distribuiscono a caso, ma secondo schemi ben stabili, almeno nelle grandi
linee, presso i pesci e gli anfibi.
Nei primi stadi di sviluppo tutti i
neuromasti sono superficiali ma in seguito possono
invaginarsi in dei canali
d’origine epidermica (pesci), mentre altri possono rimanere
superficiali (agnati
attuali).
Morfologia
Gli organi periferici del sistema laterale presentano una gran varietŕ di
struttura o di posizione; possono essere superficiali, infossati in delle
depressioni epiteliali o in dei canali; possono costituire dei bottoni ben
individuabili o al contrario delle zone estese. Due strutture distinte dei
meccano-sensori sono presenti nel sistema della linea laterale.

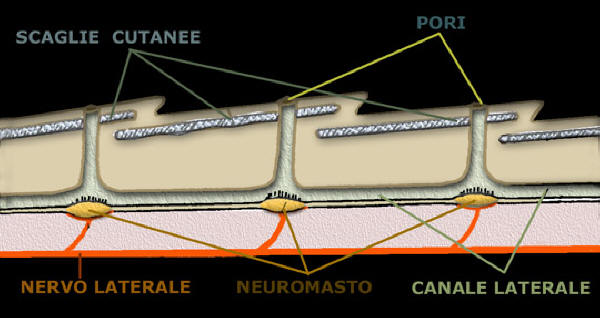
Sistema della linea laterale e sue ramificazioni in un teleosteo; suo dettaglio visto in sezione
I neuromasti liberi e i canali con neuromasti
I primi sono piccole chiazze nella pelle che sovente formano dei gruppi o linee
chiamati "Stichi", i secondi sono simili ma collegati con il fluido della linea
laterale situati sotto la pelle.
Il neuromasto
E' un organo a forma di
cupola che poggia con la sua base larga sulla membrana basale, la sua sommitŕ
raggiunge la superficie epiteliale ed entra in contatto con l'ambiente esterno:
a - Cellule di sostegno occupano tutta l'altezza dell'organo,
si assottigliano verso la superficie insinuandosi tra le cellule sensitive
degli strati superiori.
b - Cellule sensoriali d’aspetto tipicamente piriforme,
occupano lo strato superiore dell'organo. Il nucleo, sovente voluminoso, č
in posizione centrale, la zona apicale si assottigliata
si sporge
all’esterno; porta una cilia che poggia su uno stato intracellulare da
dove parte un corto prolungamento che penetra nel citoplasma.
Ciascun neuromasto contiene da alcune a qualche centinaio di cellule cigliate
generalmente con una cupola gelatinosa che include i fasci di cilia che si
estendono in alto al disopra dell’epitelio.
I neuromasti hanno le cilia
orientate in due opposte direzioni che sono completamente mescolate. Le cellule ciliate della linea laterale mostrano inoltre fascetti di cilia di
diverse dimensioni.
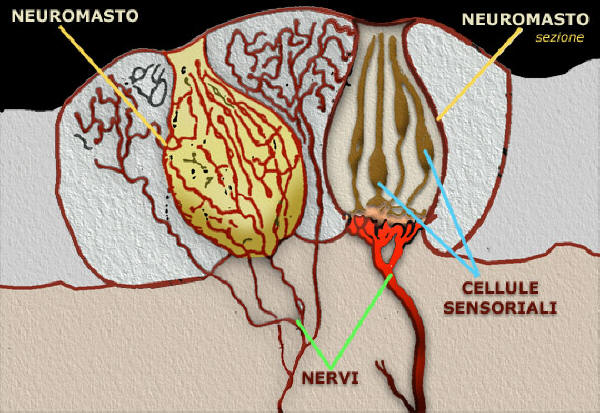
Neuromasto della linea laterale di anguilla
La struttura dei tessuti accessori č importante per il funzionamento dei
meccanosensori delle strutture della linea laterale: per esempio la forma e la
dimensione della cupola varia considerevolmente dipendendo dalla sua
collocazione del neuromasto nel corpo e anche dalla specie.
a - il neuromasto č in posizione superficiale nella pelle o
al massimo appena infossato: fossette sensoriali isolate o raggruppate
2 - il neuromasto č contenuto in un canale d'origine
epidermica
3 - il neuromasto č individualmente infossato in
un’invaginazione epiteliale che resta in comunicazione con l'esterno con un
colletto e un poro.
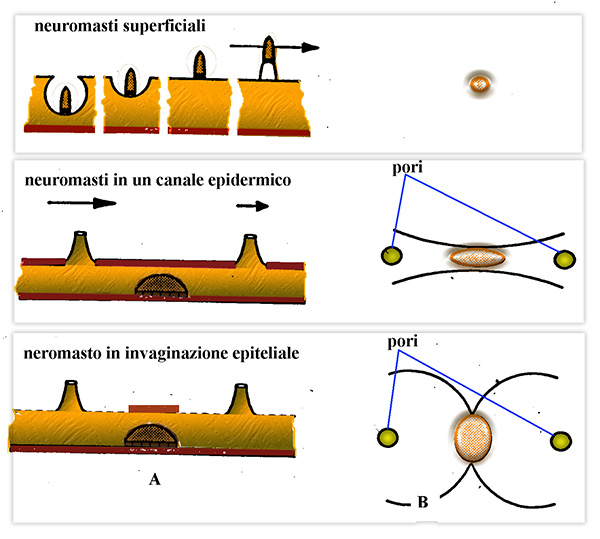
Schema delle variazioni di posizione dei neuromasti della linea
laterale
A - visti lateralmente, mostra il flusso dell’acqua, le linne marcate
indicano strutture solide mentre quelle tratteggiate strutture flessibili.
B
- la vista superiore mostra le varie forme cupole mostrate in A e la forma dei
canali e i pori
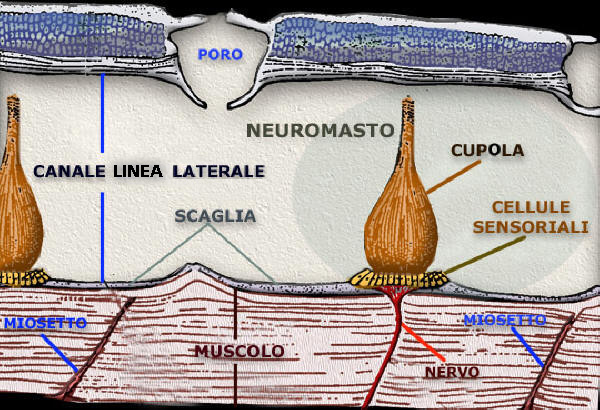
Osteitti
Il sistema laterale, innervato in prevalenza dal VII° e X° paio nn cc, č ben
sviluppato in tutti gli osteoitti e consiste nei bottoni sensoriali del tipo di
neuromasti che sono contenuti sul fondo di canali sottocutanei che si aprono
mediante pori alla superficie della pelle.
I canali sono pieni di un secreto
mucoso in cui pescano le cilia sensitive dei neuromasti. Il piů cospicuo č il
canale laterale, che percorre l'intera lunghezza del tronco e della coda su ambo
i fianchi dell'animale e che č ben riconoscibile perché di norma coperto da una
fila di scaglie cutanee laterali, ciascuna perforata da un orifizio attraverso
il quale decorre il tubulo che si apre in un poro alla superficie cutanea.
Da
ambo i lati della testa, il sistema della linea laterale si ramifica in piů
canali, dei quali i principali sono quelli delle linee mandibolari, infraorbitale e sopraorbitale; le linee sopraorbitali dei due lati sono
connesse fra loro dorsalmente da una linea commissurale.
Il sistema informa
l'animale fermo o in nuoto dello scorrere dell'acqua sopra la pelle e, in
accordo con la sensibilitŕ del labirinto membranoso dell'orecchio interno,
contribuisce a determinare il comportamento, per esempio regolando il reotropismo negativo in molti osteitti che nei fiumi usano nuotare
controcorrente.
Elasmobranchi
In essi derivano dal medesimo placoide acustico dalla cui parte mediana derivano
tanto i neuroni somatosensitivi dei gangli dell'VIII° paio di nervi cranici,
quanto l’otocisti, cioč l'abbozzo dell'epitelio che tappezza il labirinto
membranoso.
In modo analogo, da una parte delle cellule ectodermiche delle aree
cefaliche caudale del medesimo placoide derivano, rispettivamente i neuroni
somatosensitivi del VII°, del IX° e X° paio dei nervi cranici; altre cellule
delle medesime aree del placoide, migrando lungo determinate linee direttrici,
danno origine a neuromasti disseminati nella pelle della regione cefalica o del
tronco e della coda, di cui i primi innervati dal VII paio mc (nervo laterale
anteriore) e i secondi dal IX° e X° paio nn cc (nervo laterale posteriore.
Sono
allineati sul fondo di docce cutanee chiuse in forma di canali comunicanti con
l'esterno mediante pori intervallati regolarmente.
Questo sistema di canali,
ravvolti da un manicotto connettivale talora rinforzato da cartilagine, č
soprattutto sviluppato nella regione cefalica ove č suddiviso in piů rami, come il
sopraorbitale e l'infraorbitale, variamente anastomizzati e di cui il
sopraorbitale si prolunga caudalmente sotto forma di un
canale laterale che
percorre dritto in sede dorsolaterale su ambo i fianchi, il tronco e la coda
dell'animale.
Il disegno delle ramificazioni cefaliche ha subito cambiamenti nel
corso della filogenesi e pertanto ha interesse sistematico.
I neuromasti del
sistema della linea laterale fungono essenzialmente da meccanorecettori che
informano l'animale fermo o in movimento delle variazioni di pressione
dell'acqua circostante.
Negli Elasmobranchi, sono associati topograficamente
altri organi di senso simili per origine, innervazione e struttura.
Ampolla del Lorenzini
Si tratta di gruppi di minuscoli tubuli affondati nella pelle della regione
cefalica, innervati dalle terminazioni di fibre somatomotrici del VII° paio,
contenenti un fluido gelatinoso che emerge dal poro escretore per leggera
pressione su di essi esercitata. in ciascun gruppo ogni ampolla consiste in un
canale piů o meno lungo che si apre sulla superficie cutanea con un minuscolo
poro e la cui base, raggiunta dalle fibre nervose, generalmente si dilata in
diverticoli.
a - l'invaginazione, poco marcata con grande apertura
all'esterno.
b - l'invaginazione, molto marcata comunica con una stretto
colletto all'esterno , la sua estremitŕ cieca puň esser semplice o
ramificata.
c - l'invaginazione, vescicolosa č chiusa: ampolla del
Savi
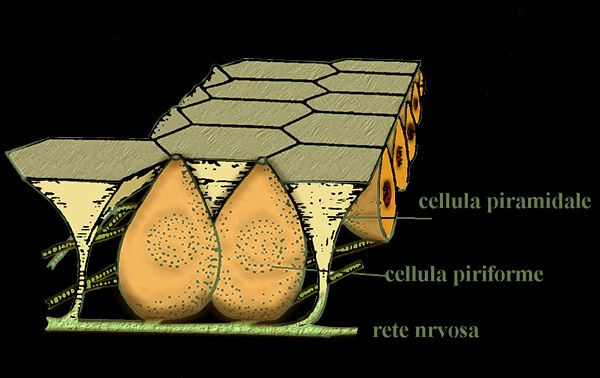
Schema dell’epitelio che riveste le ampolle di Lorenzini
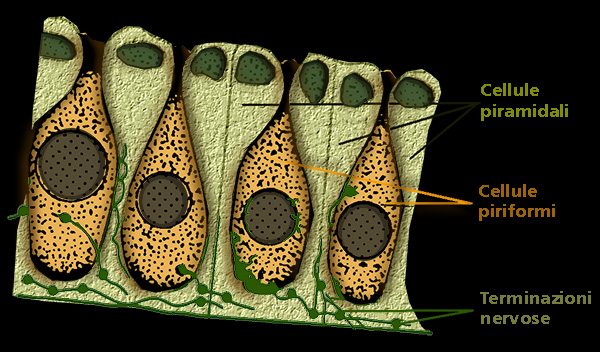
Sezione istologica
dell'epitelio che riveste le ampolle di Lorenzini
La parete del canale č tappezzata da un epitelio pavimentoso semplice; la parete
della base č invece tappezzata da uno strato di cellule piramidali a
superficie appiattita - riccamente innervate e costituenti l'epitelio sensoriale
diverso dai neuromasti perché' ogni cellula non presenta molte cilia ma molti
microvilli o talora (razze) un unico flagello - intercalate da grosse cellule
piriformi che secernono la sostanza gelatinosa contenuta nel canale.
Le ampolle
del Lorenzini vennero un tempo interpretate come organi destinati a percepire le
variazioni della pressione idrostatica tramite cambiamenti del grado di
rigonfiamento del loro contenuto gelatinoso che preme sull'epitelio sensoriale,
e dunque deputati ad informare l'animale della profonditŕ alla quale si trova,
funzione svolta in molti osteitti dalla vescica natatoria.
Studi recenti hanno
dimostrato che si tratta d’organi elettrocettori, e che hanno anche funzione
termocettrice, informando dei cambiamenti della temperatura dell'acqua.
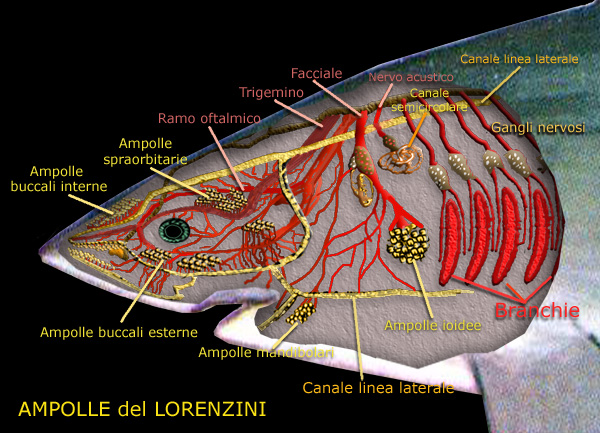
Distribuzione delle ampolle di Lorenzini, dei canali sensoriali
e relativa innervazione in una testa di squaloideo
Vescicole del Savi
Altri
organi associati al sistema cefalico della linea laterale sono in alcuni batoidei le vescicole del Savi.
Si tratta di vescicole chiuse, allineate lungo i nervi del sistema della linea
laterale specialmente nella faccia ventrale del "disco" del batoideo e innervate
ciascuna da un proprio ramuscolo nervoso.
Ogni vescicola č piena di un proprio
liquido vischioso, tappezzata da un epitelio in parte sensoriale, e provvista
alla sua base di tre dischi allineati, di cui il centrale piů
grande, ciascuno
innervato separatamente, il loro significato funzionale č incerto.
Nel loro
insieme tutte strutture sensoriali cutanee conferisce alla pelle degli Elasmobranchi la capacitŕ di percepire svariatissime stimolazioni dall'ambiente
circostante, estremamente importanti per il comportamento animale e per azioni
riflesse o volontarie.

L'Orecchio Interno
L'orecchio interno, o labirinto membranoso, deriva da una vescicola otica
dell'embrione - d’origine ectodermica (placoide uditiva) - piena di un fluido
detto endolinfa ed č compreso nel labirinto cartilagineo scavato nelle
regioni otiche del neurocranio, dove č immerso un altro liquido detto perilinfa.
L'otocisti nell'embrione si compone in una parte piů ampia detta tasca
vestibolare e una piů piccola detta tasca coclearie fra loro separate da un
restringimento che darŕ origine al labirinto composto di un utricolo e
un sacculo che un restringimento separa dalla macula della lagena e
di tre canali semicircolari (due verticali perpendicolari tra loro e
l'altro orizzontale).
Le ossa che delimitano la cavitŕ otica sono: il frontale,
il parietale, lo sfenotico, l'opistotico, l'occipitale.
Nei selaci la capsula
otica č contenuta interamente nello spessore della cartilagine.
Particolaritŕ del labirinto
Selaci
I tre canali semicircolari presentano delle disposizioni assai diverse.
Nel genere torpedo i canali verticali sono completamente indipendenti l'uno
dall'altro, ed č nell'anteriore che comunicano le estremitŕ del canale
semicircolare esterno (orizzontale).
In questo tipo di labirinto si possono
distinguere:
a -
una parte anteriore comprendente il canale semicircolare
verticale anteriore, il canale orizzontale e l'otricolo
b
- una parte posteriore con il canale verticale
posteriore e il sacculo.
Presso gli altri selaci i due canali semicircolari verticali si avvicinano al
punto che le loro estremitŕ girate verso la zona interna vengono a contatto tra
loro e sono separate da una membrana (es: leomargus) o possono fondersi insieme
(es:chimera) (che č la condizione comune a tutti i canali semicircolari
verticali dei vertebrati).
In ambedue i modelli di orecchio interno, dalla zona
di congiunzione fra utricolo e sacculo pare un canale verticale mediano: il
dotto linfatico che si apre nello spazio sottomeningeo periencefalico e
all'esterno sulla faccia dorsale del cranio, questi fa comunicare il labirinto
con l'esterno; contiene un fluido gelatinoso in cui si possono osservare delle
particole solide provenienti dall'esterno.
Pesci non selaci
Il labirinto si presenta come nella chimera ma con un canale endolinfatico in
meno; i due rami dei canali semicircolari verticali sono fusi insieme salvo nei
pesci abissali.
Le dimensioni relative dei canali variano da un genere all'altro
sono molto grandi nei mormyrili, cyclopteri.
Il sacculo si separa in due
compartimenti, uno, il piů grande, il sacculo p.d. e l'altro la lagena
(questa
č ben delimitata nell'acipenser).
Il sacculo in certi pesci č molto grande da
superare in altezza l'arco superiore dei canali verticali.(gobius niger). Manca
il canale endolinfatico o persiste sotto forma di canale cieco.
Struttura della parete del labirinto
Il rivestimento interno del labirinto č formato da un epitelio pavimentoso
semplice, si ispessisce localmente in alcune tipiche placche sensitive,
esternamente č rivestito da tessuto connettivo fibroso ben vascolarizzato.
Le
cellule sensoriali, nei canali semicircolari, si localizzano nei rigonfiamenti
detti ampolle del tratto di origine dei tre canali, a causa del loro
aspetto e della loro localizzazione sono indicate con i nomi di cresta
ampolliera anteriore, esterna o laterale e posteriore, le altre, piů
appiattite sono chiamate macule o papille.
Si ditinguono per la loro
sede, una macula dell'otricolo, una del sacculo e una della
lagena e talora una macula negletta vicino al punto in cui dal
sacculo si passa del canale semicircolare posteriore.
Le creste ampollosi
anteriore ed esterna e la macula dell'otricolo, appartenenti alla parte
anteriore dell'orecchio interno, sono innervate dal ramo anteriore del'VIII°
paio di nervi cranici.
Le macule del sacculo e della lagena, la macula negletta
e la cresta ampollare posteriore sono innervate dal ramo posteriore del medesimo
nervo.
L'epitelio sensitivo di queste strutture, stimolato dai movimenti
dell'endolinfa, informa l'animale della posizione della sua testa nello "spazio"
e dell’entitŕ delle accelerazioni o decelerazioni che si attuano durante il
movimento sia della testa sia di tutto il corpo.
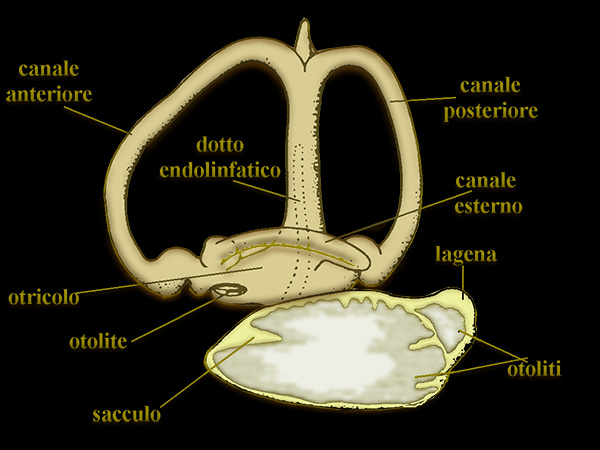
Orecchio interno ed otoliti di Teleosteo ( Perca fluvialis)
L'orecchio interno degli osteoitti ha un sacculo particolarmente sviluppato con
annessa una piccola lagena, che specialmente nei Teleostei spesso contengono otoliti assai
voluminosi, queste caratteristiche concrezioni di natura calcarea si accrescono
per opposizione superficiale in maniera discontinua, perché il loro aumento di
volume di norma si sospende nei mesi invernali.
Osservati in sezione presentano
un’alternanza di cerchi chiari e cerchi scuri attorno ad un nucleo centrale, i
cerchi chiari essendosi deposti in primavera-estate e quelli scuri
estate-autunno (valutazione dell'etŕ).
Il labirinto e il sistema laterale hanno
origine dalla medesima origine embrionale, i rispettivi recettori sensoriali
hanno delle strutture simili o identiche, infatti che nella macula, le creste
del labirinto e nei bottoni del sistema laterale i recettori sensoriali sono
fatti da uno o piů fascetti di cellule piriformi, da dove si eleva un ciuffo di
flagelli non contrattili (uno per cellula) incluso in un gel che le da la forma
di cono la cupola. Medesima importanza puň esser data alle secrezioni mucose
nell’orecchio interno, la perilinfa e vischiosa, il muco della linea laterale e
del labirinto hanno quasi la medesima composizione (mucoproteine).
Funzioni del sistema laterale
I pesci apprezzano la corrente dell'acqua grazie alla linea laterale, un oggetto
in movimento nell'acqua rompe l'equilibrio del mezzo liquido e produce anche
delle vibrazioni, la rottura dell'equilibrio causa delle correnti che esercitano
delle deboli pressioni sui recettori sensoriali, le vibrazioni meccaniche
agiscono nella medesima maniera ma le variazioni di pressione che essi creano
sono ritmiche che puň esser importante per gli stimoli deboli agendo per
sommatoria. Nei canali sottocutanei il cambiamenti di pressione si
trasmetteranno mediati dal muco.
Il sistema tattile
Il tatto, in compartecipazione dell'elettroricezione,
č un senso su cui gli squali fanno affidamento per
percepire la posizione di una preda. Proprio per questo, la
loro superficie corporea č provvista di meccanorecettori,
costituiti da terminazioni nervose libere che, oltre dagli stimoli tattili,
possono essere anche attivati da stimoli nocivi o variazioni di temperatura.
In particolare, al livello della regione superficiale, si
trovano recettori tattili ad adattamento rapido e lento, quali corpuscoli
di Meissner e dischi di Merkel.
I primi hanno la funzione di percepire gli stimoli derivanti da oggetti in
movimento che entrano in contatto con la cute, mentre i secondi sono necessari a
discriminare le caratteristiche stesse dell’oggetto.
In profonditŕ invece, in corrispondenza del derma, sono presenti ulteriori
recettori ad adattamento lento e rapido che sono, rispettivamente, i corpuscoli
di Ruffini e di Pacini. I primi, che non sono lamellari, sono deputati a
percepire lo stiramento della cute mentre i secondi, lamellari, hanno la
funzione di rispondere alle variazioni di velocitŕ degli stimoli.
Il sistema di elettroricezione
Viene
sfruttato in due modi:
- passivamente, misurando il gradiente esterno dell’acqua
che tende a fluire attraverso il campo magnetico terrestre
- attivamente, misurando i gradienti elettrici da loro
prodotti durante il nuoto all’interno del sistema terrestre.
Lo squalo bianco utilizza l’elettroricezione per captare le prede al buio e,
allo stesso tempo, puň utilizzare le ampolle per orientarsi nelle grandi
migrazioni sfruttando il campo magnetico terrestre.
Il senso dell'equilibrio
I pesci rispondono ai cambiamenti della posizione del corpo in rapporto alla
direzione della forza di gravitŕ con dei riflessi statici e dei riflessi
dinamici:
a -
I riflessi statici
consistono in delle posizioni
compensatrici degli occhi, della testa, delle pinne, del tronco in riguardo
a dei cambiamenti d'orientamento del pesci in rapporto alla verticale.
b -
I riflessi dinamici - consistono in dei movimenti
con effetto compensatorio che sono provocati da accelerazioni angolari
durante le rotazioni e da accelerazioni lineari durante le traslazioni
lineari, questi sono causati dall'azione subitanea dei muscoli effettori e
differiscono dai riflessi statici per la brevitŕ del loro periodo di
latenza.
Sembra che i riflessi statici siano la risposta all'eccitamento alla macula
dell'utricolo e i riflessi dinamici all'eccitamento delle creste dei canali
semicircolari.
Funzione degli organi otoliti nel senso dell'equilibrio
Questi organi hanno in comune le concrezioni calcaree che possono opprimere lo
strato di muco che circonda i flagelli delle cellule sensoriali.
L'otolita
esercita una duplice stimolazione:
a - preme sempre seguendo la verticale, secondo la gravitŕ,
le cupole sottostanti (funzione statica)
b - a ogni movimento del pesce, l’otolita si sposta sulla
macula e in certi punti ne eccita le cupole secondo certe direzioni
(funzione dinamica)
I movimenti sono cosi percepiti immediatamente dal pendulo a torsione dei canali
semicircolari e dagli organi a otoliti del labirinto. Gli organi a otoliti sono
sensibili anche a vibrazioni sonore (funzione acustica)
Prof. U. Fazzini
Igiene e Sanitŕ animale
Acquacoltura e Ittiopatologia
2004-2005
AA.VV.
|