|
Pesci - Anatomia - Organi Respiratori
|
|
Branchie
Osmoregolazione branchiale |
Nei processi di osmoregolazione, oltre ai reni, intervengono le branchie.
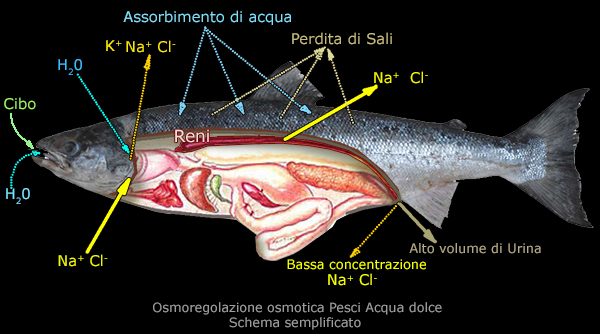
I pesci d'acqua dolce sono iperosmotici,
avendo una concentrazione
salina (Na+, Cl-) nei fluidi interni superiore a quella
dell'ambiente in cui vivono, e pertanto, quando il sangue
raggiunge le branchie, i sali tendono a diffondersi (attraverso
superfici corporee permeabili) nell'acqua, mentre l'acqua
passivamente (per
diffusione branchiale) tende ad entrare nel loro corpo diminuendo la
concentrazione salina.
Al fine di raggiungere un equilibrio osmotico interno, i pesci
d'acqua dolce rilasciano, attraverso i reni, l'acqua
in eccesso come urina diluita (che comunque contiene ioni) e
limitano l'assunzione di acqua attraverso la bocca.
La perdita dei sali avviene attraverso i filamenti branchiali e relative lamelle secondarie.
Una situazione inversa si ha nei pesci ossei iposmotici d'acqua
marina.
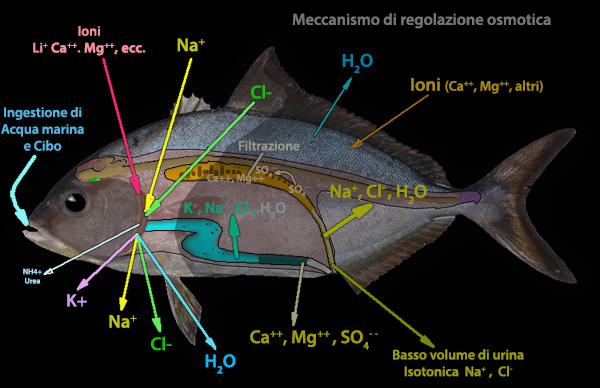
Avendo una concentrazione ipoosmotica (fluidi interni
pari ad un terzo di quella del mare), l'acqua del sangue tende a
migrare,
attraverso membrane cellulari che utilizzano proteine di
membrana chiamate acquaporine, fuori del corpo, mentre i sali
(Na+, Cl-) presenti nell'ambiente tendono ad entrare dentro,
attraverso membrane cellulari,
utilizzando dispositivi trasportatori o canali specifici attivi o
passivi.
Per evitare la disidratazione e trovare un equilibrio osmotico, i pesci
bevono molta acqua, che verr� assorbita dall'epitelio
dell'intestino. Ma insieme all'acqua entrano anche molti sali che
vanno a sommarsi a quelli assunti per diffusione dalle branchie.
I pesci cartalaginei e alcuni primitivi pesci ossei mantengono un equilibrio
osmotico differente.
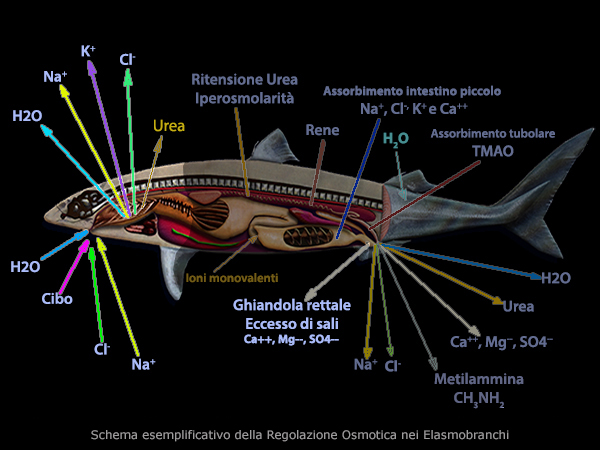
Questi pesci bevono poco e riescono a mantenere alti i levelli di
urea nel loro sangue, di conseguenza i loro fluidi interni diventano
pi� salati (iperosmotici) dell'acqua marina; un po' d'acqua salata
entra in corpo attraverso le branchie rendendo necessaria
l'escrezione dei sali attraverso una ghiandola rettale (ghiandola
del sale) che deriva dall'intestino posteriore.
Di
conseguenza l'eccesso di sali vengono escreti, per trasporto attivo
di Na+, Cl- e forse K+,
dall'epitelio delle branchie grazie alle cellule cloruro , mentre gli ioni bivalenti vengono eliminati dai reni.
Sandrine Heusser
Licence de sciences de la vie -
F.S.T Universit� Jean Monnet de
Saint-�tienne
D�partement de biologie-biochimie
2017
AA.VV.
Pesci d'acqua dolce
Ci sono state
nel tempo varie interpretazioni su come i pesci d'acqua dolce
riescano a compensare
la perdita di sale e ad invertirne il flusso dalle branchie.
Le prime ricerche hanno indicato che
gli enzimi (ATPasi) nel tessuto delle branchie dividono l'ATP e spostano
i sali contro il gradiente osmotico con l'energia che viene
rilasciata.
Si presumeva che la maggior parte di questa attivit� si
verificasse in cellule specializzate nell'epitelio branchiale chiamate
cellule beta cloruro (per differenziarle dalle cellule alfa cloruro
anatomicamente diverse presenti nei pesci di acqua salata).
Le cellule
cloruro sono ionociti
dotati di una superficie di scambio molto ampia (grazie a numerose invaginazioni della membrana
fittamente intrecciate in una rete tubulare endoplasmatica liscia) e
di
molti mitocondri che forniscono l'energia sotto forma di ATP,
che potrebbe essere utilizzata dalle ATPasi.
Questa teoria era intuitivamente attraente perch� le ATPasi potevano
scambiare Na+ con NH4+ e Cl- con HCO3-.
Lo scambio di cariche simili
avrebbe promosso l'equilibrio acido-base e sia NH4+ che HCO3- sarebbero
sottoprodotti modificati e di scarto del metabolismo.
Le cellule
cloruro, come le cellule epiteliali contengono
l'enzima anidrasi carbonica. Sembrava logico che l'anidride carbonica e
l'ammoniaca si diffondessero nella cellula cloruro dal sangue e
l'anidride carbonica, catalizzata dall'anidrasi carbonica, si sarebbe
dissociata in bicarbonato e ione idrogeno.
L'idrogenione avrebbe ionizzato l'ammoniaca
come ionio ammonio (NH4+) e la CO2 come bicarbonato (HCO3-), che
sarebbero stati escreti da Na+
-NH4+ ATPasi e Cl-
-HCO3- - ATPasi, mentre il sale sarebbe
stato pompato verso
l'interno.
Inoltre, Na+
- K+ ATPasi situata sulla membrana interna
avrebbe aiutato a spostare Na+ dal citoplasma al sangue.
Ulteriori ricerche, tuttavia, hanno indicato che erano coinvolti anche
altri enzimi e un gradiente elettrochimico.
Una teoria meno datata
sosterrebbe che, mentre tutti i movimenti sopra descritti senza dubbio si
verificano, Na+ seguirebbe anche un gradiente elettrico nella cellula
(la cellula pi� carica negativamente dell'acqua).
L'effetto combinato del
gradiente elettrico e del gradiente osmotico opposto � definito
gradiente elettrochimico, il quale sarebbe vicino a 0 per i pesci d'acqua dolce
e
bloccherebbe il deflusso di sale dalla branchia.
Questo potenziale
elettrico sarebbe mantenuto da una diversa ATPasi che pomperebbe H+ fuori dalla
cellula nell'acqua e da una Na+-K+ ATPasi che scambierebbe 3 Na+ per 2 K+
nella membrana interna.
Ma, non � chiaro quale di questi movimenti possa
verificarsi nella cellula del cloruro e quale possa essere localizzato
nelle cellule epiteliali.
La CO2 si diffonderebbe dal
sangue nella cellula del cloruro dove reagirebbe per formare HCO3-.
Infatti, la maggior parte dell'anidride carbonica si troverebbe gi� come
bicarbonato una volta raggiunta la branchia e sarebbe in grado di
modificare il pH del sangue, essendo ione predominante; la reazione da CO2 a HCO3- � catalizzata
dall'enzima anidrasi carbonica nei globuli rossi. Anche la cellula del cloruro
contiene anidrasi carbonica e sarebbe in grado di catalizzare qualsiasi CO2
rimanente in HCO3- . lo HCO3- gi�
nel sangue si diffonderebbe direttamente alla pompa ionica e verrebbe
scambiato con Cl-.
Poich� non c'� anidrasi carbonica nell'acqua, la
lenta conversione in CO2 non avvebbe finch� l'acqua non sia uscita dalla
branchia. Pertanto, la CO2 non si diffonderebbe nuovamente nel pesce.
Pesci d'acqua salata
L'attivit� ATPasi nella
branchia del pesce di mare � relativamente pi� alta, perch� il sale
deve essere pompato attivamente nell'intestino affinch� si verifichi
l'idratazione.
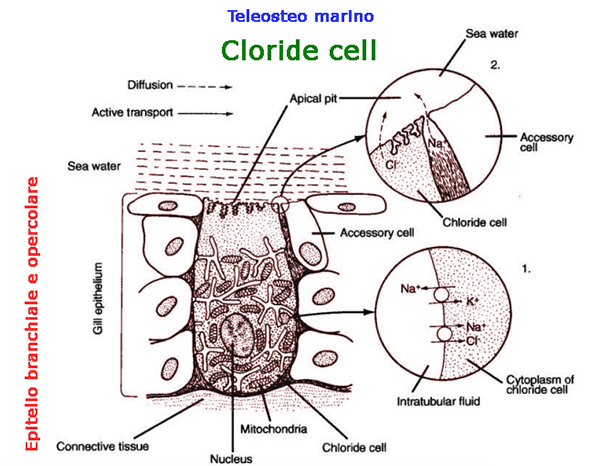
Quindi, nella branchia, i pesci
marini dovrebbero liberare il
loro corpo sia dal sale acquisito attivamente che da quello acquisito
passivamente. Inoltre, potrebbe essere necessario che i pesci
di mare impieghino il pompaggio ATPasi di Na+ in cambio di NH4+ per
liberarsi dei rifiuti azotati.
Na+-NH4+ ATPasi e
Cl--HCO3- ATPasi non
aiuterebbero l'osmoregolazione dei pesci marini come si presume facciano
con i pesci d'acqua dolce.
Si ritiene che i gradienti elettrici nella branchia del
pesce di mare spostino gli ioni necessari. In questi gradienti, l'acqua
salata sarebbe caricata negativamente, la cellula alfa del cloruro
sarebbe pi�
altamente negativa e il sangue sarebbe caricato positivamente. Il gradiente
elettrico sarebbe mantenuto da Na+-K+ ATPasi situata in un esteso sistema di
microtubuli.
Il pompaggio attivo di Na+ fuori dalla
cellula del cloruro determinerebbe un gradiente di Na+ (pi� alto nel sangue
che nella cellula del cloruro) che guiderebbe un sistema di trasporto legato
a Na+-Cl- aumentando il contenuto di Cl- della cellula e quindi l'elettronegativit�
della cellula del cloruro. La cellula del cloruro altamente negativa farebbe
s� che Cl- si sposti verso l'acqua di mare meno carica negativamente
lungo il gradiente elettrico.
Il Na+ caricato positivamente seguirebbe il
gradiente dal sangue caricato positivamente all'acqua di mare caricata
negativamente all'esterno della cellula del cloruro.
Poich� questo
meccanismo di spostamento degli ioni sarebbe alimentato interamente dalla
Na+-K+ ATPasi, questo enzima aumenterebbe drasticamente quando
i pesci eurialini dovessero spostarsi dall'acqua dolce a quella salata.
L'ingestione di acqua salata non idrater� il pesce a meno che
non abbia un meccanismo per spostare le molecole d'acqua dall'intestino
al sangue contro un gradiente osmotico. Pertanto, i sali devono
essere spostati attraverso la creazione di un gradiente osmotico che
l'acqua seguir�.
Le cellule del rivestimento intestinale nei pesci
marini
usano la Na+-K+ ATPasi per creare un gradiente elettrochimico simile
alle cellule del cloruro della branchia (ma nella direzione opposta) che
spostano NaCl nel tessuto che circonda l'intestino, creando un'area
localizzata in cui l'osmolarit� � maggiore dell'acqua di mare che fa s�
che l'acqua scorra nel sangue del pesce.
AA.VV.
Recentemente nuovi studi approfonditi
hanno rimodulato i meccanismi della osmoregolazione branchiale.
Pertanto si rimanda a letture scientifiche pi� idonee.
Alcune letture di pubblicazioni indicate nei links potrebbero gi� dare
una visione pi� approfondita dei meccanismi di osmoregolazione
branchiale.