|
|
|
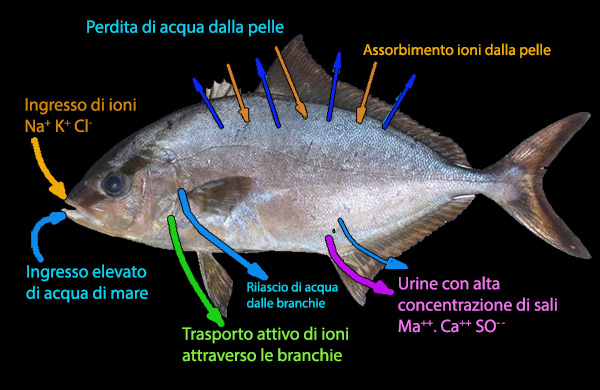
Morfologicamente, i teleostei sono ben adattati allo scambio ionico utilizzando
meccanismi attivi e passivi attraverso varie membrane
superficiali per raggiungere un corretto equilibrio osmotico e stato di idratazione.
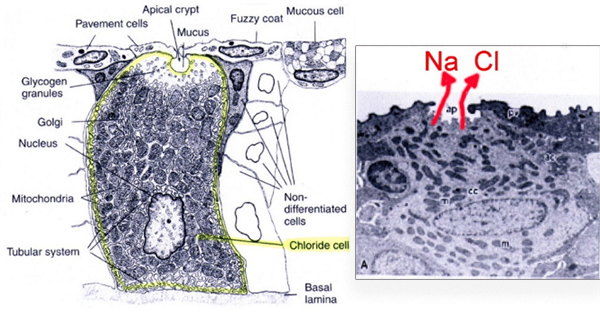
Alla base delle lamelle branchiali secondarie si possono riconoscere delle
cellule epiteliali
(cellule del cloruro, estremamente ricche di mitocondri,
polarizzate in due parti, una delle quali interagisce con l’acqua e
l’altra con il capillare sanguigno) che sono deputate al
rilascio dell'eccesso di sali, in particolare
di cloruri, proveniente dal cibo e dall'acqua ingerita. La secrezione di NaCl avviene
attraverso un trasporto secretorio primario di Cl- e un trasporto
passivo secondario di Na+.
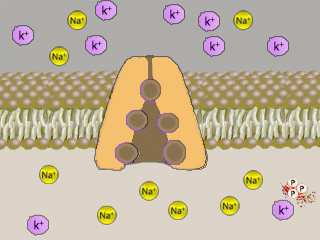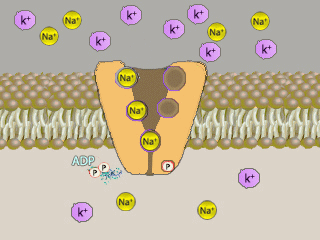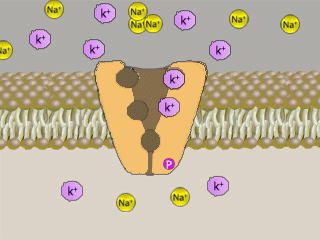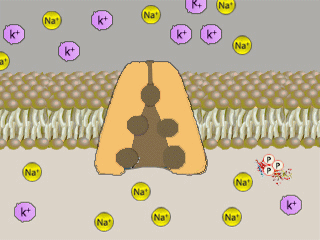
Na+K+ATPasi (NKA, conosciuta come pompa sodio-potassio) é un enzima,
localizzato in tutte le membrane basolaterali degli
ionociti e nella maggior parte delle cellule) che
svolge il ruolo di
trasportatore attivo
di sostanze nella cellula, grazie
all'utilizzo di energia.
Il meccanismo di trasporto č influenzato dalle concentrazioni di
ioni sodio e potassio:
un’alta concentrazione di Na+
porta ad un trasporto dello stesso ione fuori della
cellula, mentre un’alta concentrazione di
K+
attiva il meccanismo
contrario.
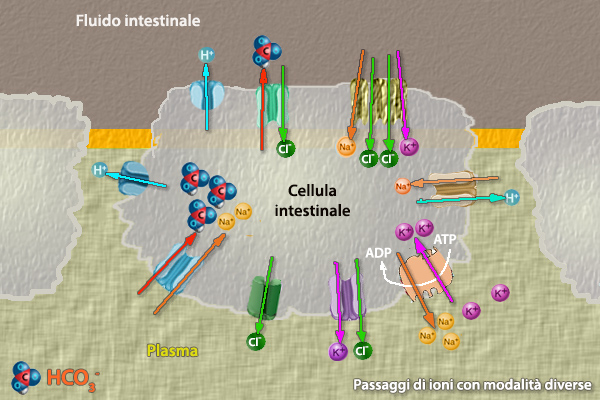
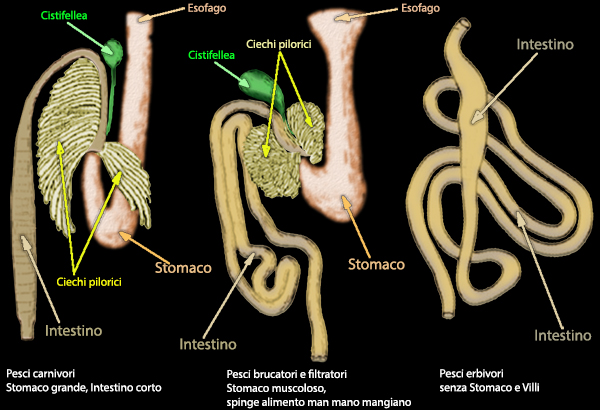
Il Tratto gastro intestinale ha la funzione di immettere nel
sangue, attraverso gli strati di cellule epiteliali altamente
polarizzate che formano la mucosa intestinale, i nutrienti provenienti dall’ambiente. Le molecole complesse vengono scomposte
mediante degradazione meccanica ed enzimatica
(proteasi, lipasi,
carboidrati, ecc.), diventando molecole semplici.
Nel caso dell'assorbimento dei lipidi dopo la lipolisi, i composti risultanti
(monogliceridi, acidi
grassi a catena lunga, fosfolipidi, colesterolo, vitamine liposolubili), formano micelle con sali biliari,
che vengono poi trasportate negli enterociti, dove vengono
riesterificate in trigliceridi. Successivamente, quest'ultimi
sono accompagnati da un numero elevato di molecole d'acqua, il cui
numero dipende dal trasportatore specifico.
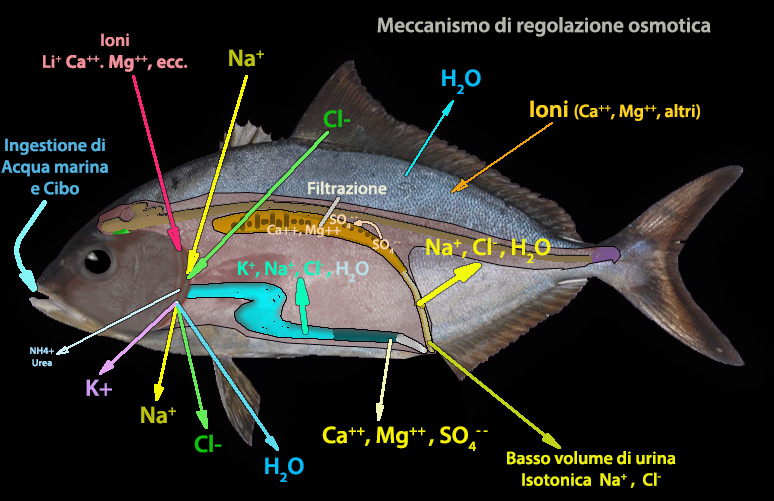
Le cellule
del rivestimento intestinale nei pesci marini utilizzano (in
direzione opposta) Na+
-K+ ATPasi per spostare NaCl nel
tessuto circostante l'intestino; viene cosě creata un'area, con osmoalitŕ maggiore dell'acqua di mare,
che favorisce lo
scorrimento dell'acqua nel sangue. La fase finale di questa idratazione č
l'escrezione del sale da parte della cellula cloruro della branchia.
L'intestino prossimale
(esofago e intestino anteriore)
rimuove fino all'80% dei principali ioni monovalenti (Na+, Cl- e K+)
assunti, mediante
trasportatori passivi che agiscono lungo un gradiente
elettrochimico e
non richiedono energia. Il fluido intestinale
elaborato, perň, diventa ricco di ioni bivalenti (Ca++, Mg++, SO4--).
Il fegato nei teleostei č una ghiandola abbastanza grande con
labatura variabile (unico lobo, bilobato,
trilobato, polilobato) e svolge la funzione di produrre
bile e sali biliari, che cooperano con gli enzimi pancreatici a
tagliare e ad assorbire i grassi. La cistifellea puň essere sferica,
ovoidale, allungata).
Recettori cationici polivalenti (della famiglia dei chemocettori di
feromoni e degli odori) sono presenti sulla superficie corporea, sul cervello, sulle
branchie, sull’intestino e sulla pelle.
Poiché
l'acqua di mare
contiene alte concentrazioni di Ca++ e Mg++, č
ragionevole ipotizzare che i recettori cationici polivalenti
extracellulari servano per l'osmosi.
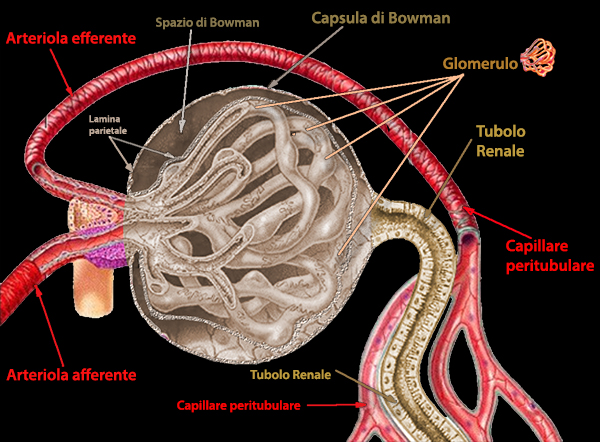
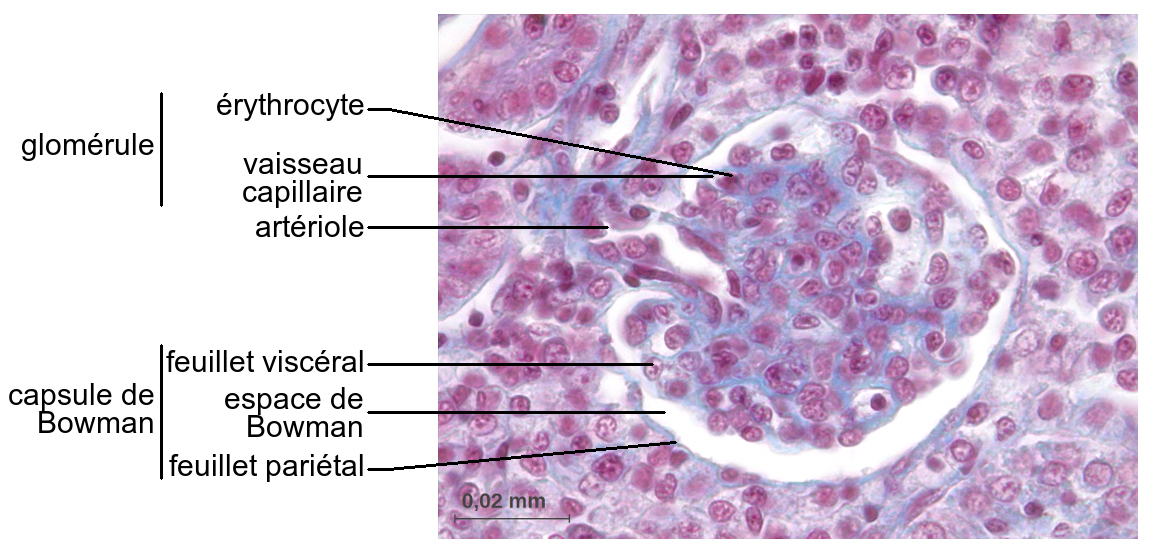
La
capsula di Bowman č delimitata esternamente da uno strato
parietale (epitelio semplice e squamoso), e internamente da uno
strato viscerale (cellule che sviluppano espansioni
ramificate). Tra gli strati parietale e viscerale si genera lo
spazio di Bowman, preposto al contenimento dell'urina. La
capsula di Bowman circonda un grappolo di vasi sanguigni capillari
(glomerulo), costituenti il corpuscolo malpighiano. Il sangue
viene portato al glomerulo da un'arteriola afferente e
ripreso da un'arteriola efferente. Nel corpuscolo malpighiano
le strutture escretrici, rappresentate dalla capsula di Bowman del
nefrone, e le strutture circolatorie, corrispondenti ai vasi
capillari del glomerulo, sono strettamente associate.
Acqua, sali e prodotti di scarto del flusso sanguigno entrano nella capsula di Bowman e poi fluiscono attraverso il tubulo renale e il dotto collettore fino al dotto urinario per essere espulsi. Attraversando il tubulo, il glucosio, diversi minerali e in alcuni casi, acqua, vengono riassorbiti. Il filtraggio e il riassorbimento sono controllati dall'azione ormonale.
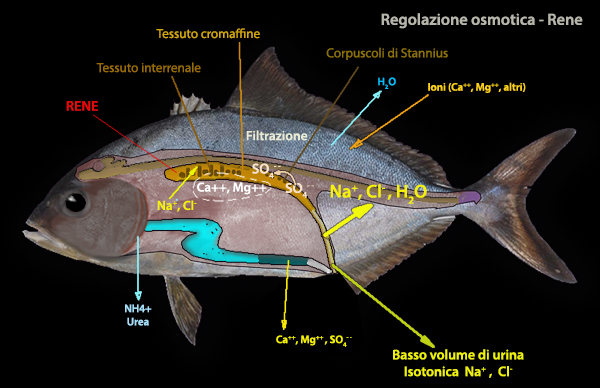
I reni sono preposte a ritenere o ad eliminare sostanze; la principale escrezione riguarda gli ioni
bivalenti. Quasi tutti
gli ioni SO4--, nella filtrazione glomerulare vengono riassorbiti dai
tubuli prossimali, (in quest'ultimi SO4-- non
innesca il passaggio dall'assorbimento alla secrezione). Diversi studi indicano che il Ca++ puň essere regolato in
differenti tessuti; Mg++ e SO4-- sembrano essere
trattati
principalmente dal rene.
Il NaCl viene per
lo piů riassorbito tramite Na+K+2Cl-.
I corticosteroidi (specialmente il cortisolo) mčdiano molte funzioni corporee dell’osmoregolazione,
quali la regolazione naturale
dell'equilibrio di ioni e acqua, la facilitazione dell'escrezione di Na+
(aumentando l'attivitŕ dell'ATPasi
Na+-K+) nelle branchie, il miglioramento e
l'assorbimento di acqua nell'intestino e nella vescica urinaria, la
risposta allo stress. Gli ormoni
dello stress svolgono un ruolo fondamentale nel mantenimento dell’omeostasi
interna, attraverso la funzione mineralcorticoide
(11-desossicorticosterone) che controlla il
trasporto di ioni e acqua.
|

|
|
|
|
|
|
|
|
|
|
|
|
|
|
Osmoregulation in fish Mechanisms and clinical implications - Martin G. Greenwell, Johanna Sherrill, Leigh A. Clayton - The Veterinary clinics - WBS -- 2003 |
|
|
OSMOREGULATORY ROLE OF THE INTESTINE IN FISH PHYSIOLOGY. - EVOLUTIONARY, ENVIRONMENTAL, AND TOXICANT IMPLICATIONS - A Ph.D Thesis by ANDRÉ BÁRANY RUIZ - University of Cádiz, Spain - February 2021 - pdf |
|
|
Compartiments hydriques et osmorégulation - Michel Tsimaratos, Laurence Dubourg - DIU de Néphrologie PédiatriqueC1 - 2019 - pdf |
|
|
Réponse au Stress Osmotique - Dr. R. ALATOU - Université Frčres Mentouri Constantine - Faculté des sciences de la nature et de la vie Département de Microbiologie - pd |
|
|
|
|
|
Variabilitŕ intraspecifica e trade-offs nel foraggiamento e nel comportamento anti-predatorio di Liza aurata (Cefalo dorato) - Fabio Antognarelli - UNIVERSITŔ DEGLI STUDI DELLA TUSCIA DI!VITERBO - DIPARTIMENTO DI ECOLOGIA E SVILUPPO ECONOMICO SOSTENIBILE - 2012 |
|
|
A brief history of the study of fish osmoregulation: the central role of the Mt. Desert Island Biological Laboratory - David H. Evans - Frontiers in physiology - 2010 - pdf |
|
|
Apparato escretore - Bertone - Lez. 27& 28 - pdf |
|
|
Fisiologia applicata Specie ittiche - Part. 1 Osmoregolazione - Parmeggiani - Seminario |
|
|
Trasporti di membrana - I. tomassini Barbarossa - Universitŕ degli studi di Cagliari - Lezioni di fisiologia |

















|
|
|
|
www.colapisci.it |
|