|
Pesci
- Anatomia - Apparato circolatorio
|
|
Osmoregolazione Elasmobranchi
|
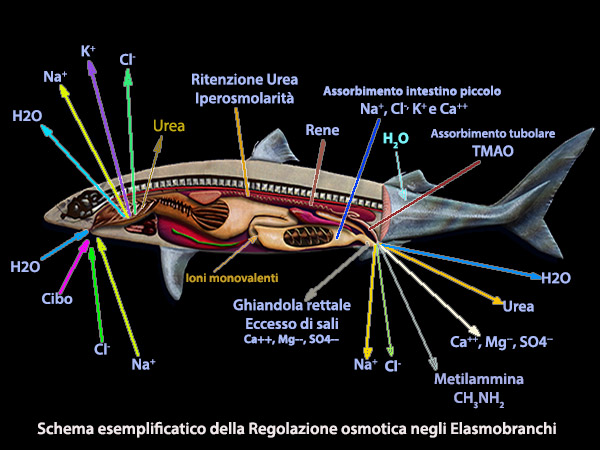
Gli elasmobranchi marini
risolvono il problema dell'osmoregolazione
divenendo leggermente ipertonici
o isoosmotici
rispetto all'acqua di mare. Infatti la concentrazione di elettroliti
del plasma č tenuta bassa. Gli elettroliti sono sostituite da molecole organiche (es. Urea)
che contribuiscono ad avere una concentrazione interna piů alta del
normale, ma che viene ben tollerata.
Le branchie degli elasmobranchi
tendono ad acquisire acqua per osmosi e i sali per diffusione. Ogni
branchia contiene due varianti morfologiche di una cellula ricca di
mitocondri; queste cellule differiscono strutturalmente
dalle cellule del cloruro dei teleostei marini e le loro funzioni
non sono ancora chiarite.
La fonte primaria di
afflusso di Na+, oltre all'assorbimento epiteliale delle branchie
e alla ingestione di acqua marina, č data dal cibo ingerito.
Le concentrazioni di ioni sodio e ioni
cloruro, superiori a quella del
mare, possono spostarsi tramite il sottile
epitelio branchiale e possono essere escrete o assorbite.
In preparazioni branchiali perfuse č stato
verificato efflusso di ammoniaca mediato dal trasportatore Na+-NH4+-2Cl-.
Questo trasportatore potrebbe fornire una via di escrezione di Na+.
L'eccesso di NaCl viene escreto, con
velocitŕ controllata da fattori umorali e neuro trasmettitori, a concentrazioni superiori a quelle del plasma, attraverso una struttura specializzata nota come ghiandola rettale. Questa ghiandola,
detta anche ghiandola del sale, č un prolungamento a forma di dito della regione colorettale.
Il sodio escreto dalla ghiandola rettale é, in percentuale,
inferiore di quello escreto dalle branchie.
Altri sali
assorbiti in eccesso vengono escreti con l'urina, specie per
gli ioni bivalenti (Ca++, Mg++ e So4--).
L'angiotensina II e l'espansione del volume sembra essere uno stimolo
importante per l'escrezione di NaCl, grazie anche alla
collaborazione del
tubulo renale, il quale assorbe l'ossido ammina di trimetile (TMAO)
(scarto azotato).
L'afflusso di urea
e TMAO, circa il 40%
dei soluti plasmatici, aumentano la pressione osmotica del sangue a un livello appena
superiore a quello dell'acqua di mare.
La pelle assorbe parzialmente acqua ed č quasi impermeabile impermeabile all'urea e al TMAO.
Gli elasmobranchi marini devono affrontare una potenziale ipervolemia
(eccessivo accumulo di liquidi). Per far fronte a questo carico d'acqua,
la velocitŕ di filtrazione glomerulare (GFR)
e i flussi di urina si avvicinano a quelli dei teleostei d'acqua
dolce.
Come i teleostei marini, possiedono meno NaCl nei loro fluidi corporei rispetto all'acqua di mare
(NaCl circa il 50% in meno).
A livello del piccolo intestino si ha invece un
assorbimento attivo
di ioni monovalenti, attraverso un cotrasportatore Na+
mentre gli ioni bivalenti Ca++ Mg++ e
SO4-- sono escreti insieme all’acqua attraverso l’ano.
Infine l’eccesso di Na+ e
parte del K+ viene eliminato dal sangue attraverso un
altro meccanismo di trasporto attivo a livello delle branchie.
Gli elasmobranchi marini mantengono un plasma iperosmotico all'acqua di mare perché mantengono
concentrazioni
molto elevate di urea (2,5%) e altri prodotti di scarto azotati nel loro sangue.
L'urea, principale prodotto finale del metabolismo proteico č molto piů stabile nel
plasma dell'ammoniaca, ma denatura le proteine.
Gli elasmobranchi marini sono in grado di tollerare elevati livelli di urea grazie agli effetti protettivi dei composti di metilammina,
come l'ossido di trimetilammina, betaina e sarcosina. La maggiore
presenza di urea rispetto all'ossido di trimetilammina riduce al
minimo la denaturazione delle proteine. Le concentrazioni di urea sembrano essere simili sia nel plasma
sia nelle cellule.
L'elevato livello di urea plasmatica riscontrato negli elasmobranchi
marini sembra essere mantenuto e regolato principalmente dal rene,
sebbene sia prodotto nel fegato.
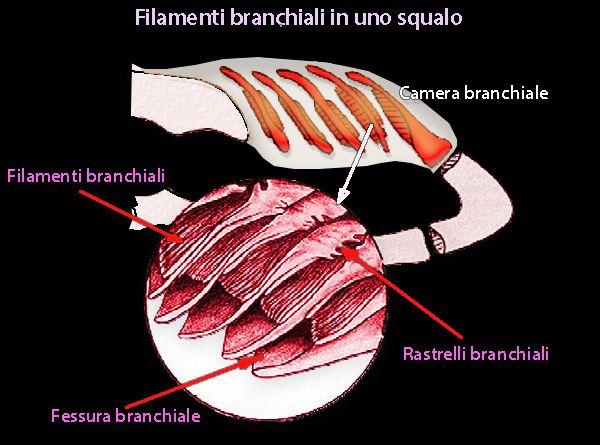
Le branchie sembrano essere il sito principale di efflusso di urea, perché le perdite renali sono solo il 4-5% della perdita totale e meno
del 15% dell'urea filtrata viene escreta. Infatti, si pensa che quasi tutta l'urea filtrata dai glomeruli venga riassorbita tramite
un sistema di scambio controcorrente all'interno del rene.
I nefroni degli elasmobranchi hanno glomerulo, collo, segmenti del tubulo prossimale I e II,
tubulo distale e dotto collettore. La disposizione dei nefroni negli elasmobranchi č
piů complessa rispetto ai teleostei. Il rene degli elasmobranchi č composto da guaine peritubulari di cellule squamose
che circondano sistemi controcorrente di diversi nefroni raggruppati insieme con anse capillari anastomizzate.
Come indicato in precedenza, per contrastare l'afflusso di acqua attraverso le branchie, il GFR č relativamente alto (400 lL/100 g/h),
come nei teleostei d'acqua dolce.
A differenza dei teleostei d'acqua dolce, tuttavia, gli elasmobranchi marini possono raccogliere sali
dal loro ambiente di acqua di mare.
Na+, Cl-, K+ e Ca++ vengono
riassorbiti dal filtrato glomerulare con rapporti urina-plasma prossimi a 1,0 per Na+ e Cl-.
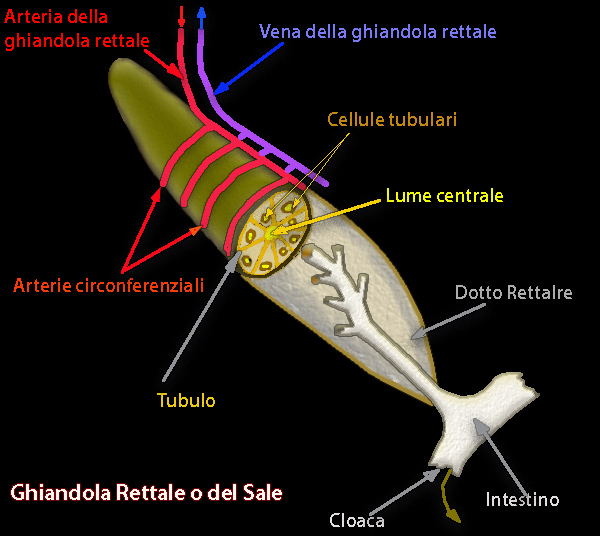
La secrezione di sale da parte della ghiandola rettale sembra essere controllata da una varietŕ di ormoni e neurotrasmettitori.
Lo
stimolante primario potrebbe essere un peptide intestinale vasoattivo o un altro peptide dell'intestino, chiamato rettina. L'adenosina
agisce come stimolante per la ghiandola rettale a concentrazioni superiori a 10-5 M.
A concentrazioni inferiori, l'adenosina agisce come inibitore della ghiandola rettale. La somatostatina
inibisce l'azione stimolatoria sia del peptide intestinale vasoattivo che dell'adenosina. L'angiotensina II sembra essere
un vasocostrittore negli elasmobranchi facilitando il rilascio di catecolamine nella periferia.
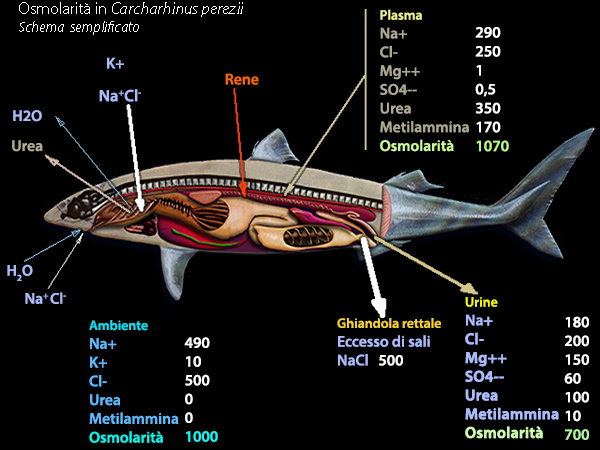
"I
Selaci marini si evolvono in un ambiente ad elevata concentrazione
osmotica, per la presenza di ioni ad elevate concentrazioni. Le
differenze di concentrazione esistenti tra l'ambiente di vita e
l'ambiente interno sono causa di flussi
spontanei di acqua e ioni a livello delle superfici corporee
permeabili come le branchie, il tegumento e il tubo digerente.
Il problema dell'acqua č in gran parte risolto dall'isoosmolaritŕ
dell'ambiente interno rispetto all'ambiente esterno. La
concentrazione osmotica dell'ambiente interno č legata alla presenza
di ioni sodio e cloruro, ma soprattutto di urea. L'urea
consente all'ambiente interno dei Selaci marini di essere in
equilibrio con l'ambiente di vita da un punto di vista osmotico. Gli
scambi idrici attraverso il tegumento sono bidirezionali e di pari
importanza, garantendo il rinnovo dell'acqua nell'ambiente interno.
Vari organi e processi, compresi i trasporti attivi, sono coinvolti
nell'osmoregolazione e nella ionoregolazione, come il riassorbimento
controcorrente dell'urea da parte del rene, la secrezione attiva di
ioni cloruro e sodio da parte della ghiandola rettale. Le
concentrazioni dell'ambiente interno in ioni sodio, cloruri e urea
vengono cosě stabilizzate e l'organismo si adatta all'ambiente
marino"
(.Le problčme de l'eau et des ions chez les Sélaciens marins
- ens-lyon.fr)
-
(Petites questions de physiologie animale (ens-lyon.fr)